
台湾泥鳅幼鱼耐盐特性研究
2022-03-11 21:30:10苗德霞陈嘉豪闫密王建华李雪隆小华朱明
江苏农业科学 2022年4期
苗德霞 陈嘉豪 闫密 王建华 李雪 隆小华 朱明



摘要:研究了盐度对台湾泥鳅幼鱼的耐受性、生长存活及呼吸代谢的影响。试验采用生态急性毒理学方法,确定试验鱼在24、48、72、96 h的半致死浓度(LC50)值分别为17.156‰、16.087‰、15.541‰和15.109‰。并以此为依据将生长和呼吸代谢实验的盐度梯度设置为0、3‰、6‰、9‰和12‰,研究幼鱼在不同盐度水体中养殖60 d生长情况、6 h耗氧率及72 h排氨率变化情况。结果表明,随着盐度的升高,台湾泥鳅幼鱼的特定生长率、耗氧率和排氨率的变化规律具有相似性,其中,幼鱼特定生长率从高到低的盐度组依次为:3‰>0>6‰>9‰>12‰;耗氧率峰值在试验开始时出现在盐度3‰组,24 h时出现在盐度6‰组;而排氨率峰值以盐度3‰组为最高。综上提示,台湾泥鳅具有一定的盐度耐受性,盐度6‰以内适宜其生长,而盐度3‰显著促进其生长和代谢。因此,在泥鳅苗种培育阶段适当提高养殖水体盐度,能够有效提高幼鱼的成活率和增长率,从而提高台湾泥鳅的养殖产量和经济效益。
关键词:台湾泥鳅;盐度;耐受性;特定生长率;耗氧率;排氨率
中图分类号:S966.4 文献标志码: A
文章编号:1002-1302(2022)04-0138-05
收稿日期:2021-06-07
基金项目:江苏省重点研发计划(现代农业)项目(编号:BE2018387);桂林市科学研究与技术开发计划(重大专项)(编号:20190102)。
作者简介:苗德霞(1980—),女,江苏连云港人,硕士研究生,主要从事水产动物增养殖研究。E-mail:64907668@qq.com。
通信作者:朱 明,博士,教授,主要从事水产生态学研究,E-mail:zhuming2382@163.com;隆小华,博士,副教授,主要从事盐碱地改造与开发利用研究,E-mail:longxiaohua@njau.edu.cn。
台湾泥鳅属鲤形目、鳅科、副泥鳅属[1],多分布于长江中下游、台湾西北部的浅滩河流中[2-3]。因其口感好、风味佳、生长速度快、对环境适应能力强且兼具药食两用价值,而受到广大养殖户和消费者的青睐。目前,对台湾泥鳅的研究主要集中在养殖与育苗[4-5]、人工繁殖和早期发育[6]、营养成分[7-9]、系统地理[10]、形态与基因遗传[2]以及生长差异性[1]等方面。
研究表明,在适宜的盐度范围内,淡水鱼类能够获得较好的生长性能,且生长率较高[11]。但当盐度超过该范围时,就会对鱼类的生理机能产生一定的负面影响。目前,有关鱼类对盐度耐受性研究主要有草鱼、松浦镜鲤、黑龙江野鲫、鲢鱼、大鳞鲃[12]、澎泽鲫[13]等。而关于台湾泥鳅对盐度耐受能力及生长和呼吸代谢等方面的影响尚未见报道。本研究首次探讨了台湾泥鳅幼鱼对盐度的耐受特性,旨在为台湾泥鳅养殖生产提供理论依据和参考。
1 材料与方法
1.1 试验材料
试验于2020年5—8月,在连云港市赣榆区丰瑞泥鳅养殖专业合作社进行。所用台湾泥鳅幼鱼由合作社人工繁殖获得,选取规格相似、游泳正常且健康的个体,放在室内暂养2周,期间投喂饲料,所产生的残饵等及时清理。试验前24 h,停止投喂。
1.2 试验条件
将速溶海水晶溶于自来水,配制成盐度为30‰的母液,于试验前24 h调配成试验所需浓度的盐溶液备用。试验期间,水温保持在(25±2) ℃,溶解氧不低于6.5 mg/L。
1.3 试验方法
1.3.1 急性毒性试验 根据预试验结果,按照等对数间距法[14]设置12.6‰、13.3‰、13.8‰、14.4‰、15.1‰、15.8‰、16.6‰、17.4‰共8个盐度组和1个对照组(0),每组3个平行,每个平行放入10尾鱼,观察并准确记录24、48、72、96 h时幼鱼的活动情况,死亡个体及时捞出。
1.3.2 生长试验 各试验组盐度分别为0、3‰、6‰、9‰和12‰,将试验鱼放入相应盐度的玻璃缸中,坚持每日08:00、17:00按时饱食投喂饲料,连续投喂60 d。
1.3.3 呼吸代谢试验 盐度梯度设置为0、3‰、6‰、9‰、12‰。耗氧试验选用3 L锥形瓶作为容器,将幼鱼放入相应盐度水体后快速封住瓶口。观察记录幼鱼的活动情况,并用溶氧仪监测溶解氧含量,试验为期6 h。排氨试验在透明玻璃缸中进行,共持续72 h,期间不投食不换水,分别于6、12、24、48、72 h时取水样,用次溴酸盐氧化法[15]检测氨氮含量。
1.4 数据处理
试验数据采用Excel办公软件进行整理,急性毒性试验结果运用改良寇氏法[16]进行分析计算。其他数据采用Origin 2018软件进行分析。差异性效果使用one-way ANOVA和LSD进行分析。
2 结果与分析
2.1 台湾泥鳅对盐度的耐受性
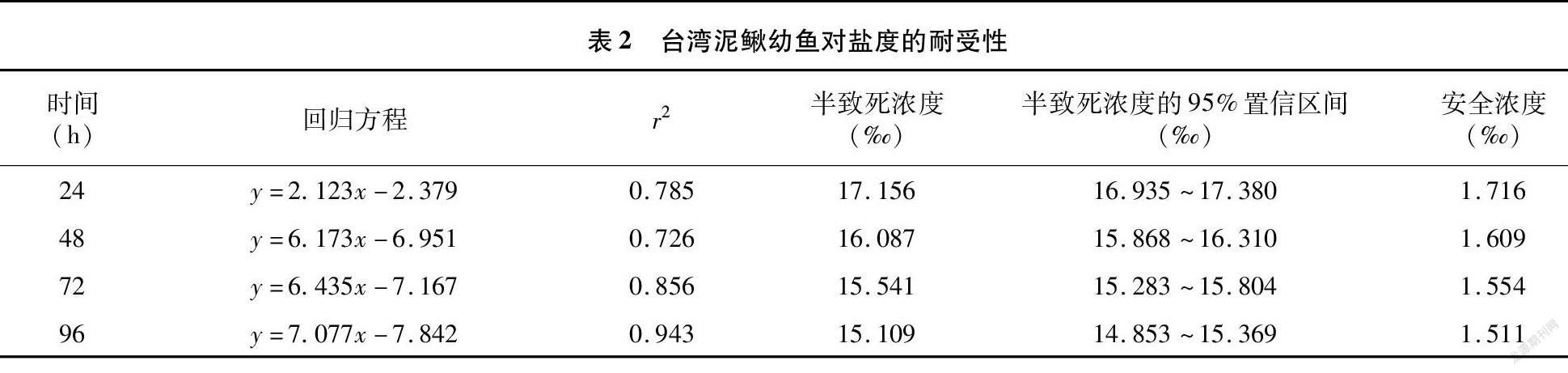
由表1可知,将幼鱼从淡水直接放入相应盐度的水体后,各试验组死亡率因盐度不同而有所差异。其中,对照组和盐度12.6‰组96 h时死亡率为0,其他各组均有死亡现象发生,且随着时间延长和盐度增加呈上升趋势。盐度15.8‰组96 h的死亡率为63.33%,而盐度16.6‰组在48 h时即达到该水平,之后缓慢上升;盐度17.4‰组在48 h时全部死亡。提示盐度对幼鱼的毒性效应与盐溶液浓度和暴露时间呈正相关,盐度越高死亡率越高,暴露时间越长死亡率越高。
由表1、表2经计算得出幼鱼24、48、72、96 h的半致死浓度(LC50)和对应的安全浓度(SC)。
2.2 盐度对台湾泥鳅幼鱼生长的影响
在不同盐度水体中养殖60 d后,台湾泥鳅幼魚的生长性能见表3。由表3可知,随着盐度增加,以终末体质量、特定生长率表示的生长性能呈先升高后降低的变化趋势。其中,幼鱼终末体质量最大值出现在盐度3‰组,为(6.955±2.436) g/尾,最小值为(2.977±0.536) g/尾,出现在盐度12‰组;特定生长率峰值也出现在盐度3‰组,且与淡水组相比差异不显著(P>0.05),但与盐度6‰、9‰、12‰组相比显著升高(P<0.05);成活率最大值为100%,出现在盐度3‰、6‰组,盐度12‰组成活率最低。由此可知,盐度3‰和6‰组的幼鱼,存活率最高,生长性能较好。
2.3 盐度对台湾泥鳅耗氧率的影响
由图1可知,在试验盐度范围内,盐度3‰组幼鱼的耗氧率在1 h时值最高,1 d时即恢复到对照组水平;盐度6‰组的耗氧率水平总体高于其他各试验组,且随着时间的延长先升高后趋于平稳,并在 4 h 时达到峰值;而高于9‰盐度组幼鱼的耗氧率一直处于较低水平,且变化不明显。
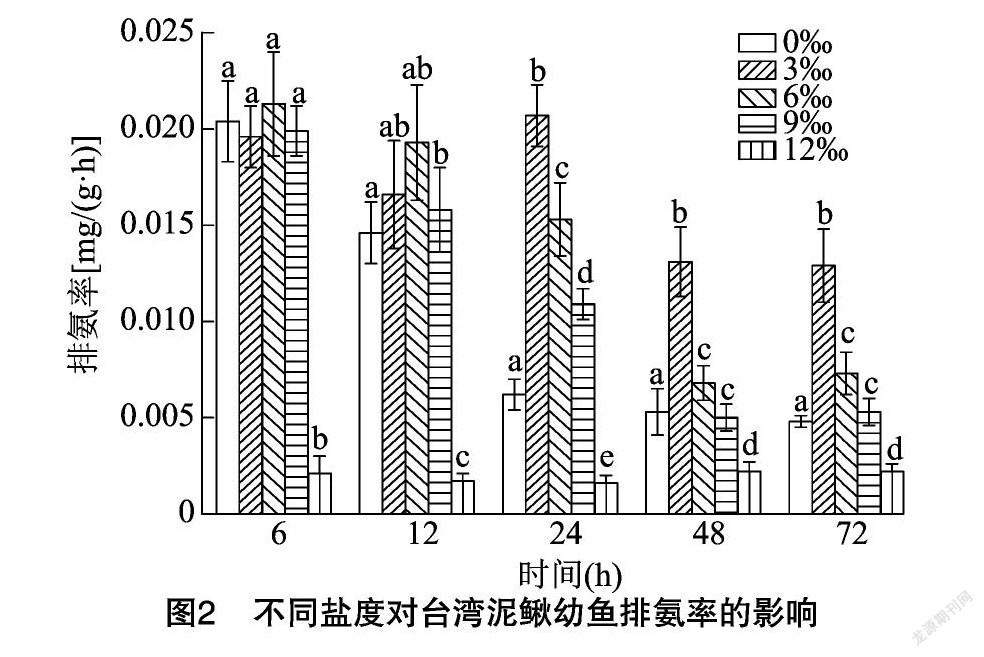
2.4 盐度对台湾泥鳅排氨率的影响
由图2可知,随着试验时间推移,盐度3‰组幼鱼的排氨率变化趋势为先升后降再升再降的变化趋势,并在24 h时取得最大值;盐度6‰、9‰组均在6 h时取得最大值,此后缓慢降低;盐度12‰组则变化不明显。
3 讨论
3.1 台湾泥鳅幼鱼对盐度的耐受性
武鹏飞等研究发现,当盐度超过3种鳅科鱼类的耐受极限值时,会加速泥鳅的死亡,呈现“阈值型”的规律[17]。与之有相似规律的还有大鳞鲃[18]、澎泽鲫[13]、花鲈幼鱼[19]、塔里木河叶尔羌高原鳅[20]等。本研究中,盐度16.6‰组幼鱼在48 h时死亡率骤然上升,盐度17.4‰组在48 h时全部死亡的现象与上述研究中观察到的结果一致,提示台湾泥鳅幼鱼调节和维持渗透平衡的能力是有限,如果水体盐度波动较大,远高于其自身对渗透压的调节能力,就会引起其短时间内快速死亡。
而盐度15.8‰组幼鱼在96h时死亡率上升到63.33%,则说明该盐度可能是幼鱼的敏感盐度,因此推测15.8‰以上盐度超出了幼鱼的渗透调节的能力范围;其余各盐度组幼鱼的死亡率随时间的推移缓慢上升,则表明其在受到盐度胁迫后,机体若不能及时启动渗透压调节机制,机体的渗透压平衡将会被破坏,进而造成细胞形态、结构和功能出现异常,将迅速死亡[21]。但随着试验持续进行和鱼体受高盐度胁迫时间的继续,幼鱼对外界高盐度胁迫的适应机制被启动以应对这种不适的环境变化情况,慢慢的有些幼鱼对这种高盐度胁迫产生了适应机制,能够逐渐适应在高盐度水体中生活,其机体内部环境也慢慢恢复到接近胁迫之前的水平,这也为解释试验结束时有些盐度组中仍有个别幼鱼存活提供了依据[21]。
3.2 盐度对台湾泥鳅幼鱼生长的影响
研究表明,淡水鱼类的生长活动受盐度影响作用明显,甚至在一定盐度范围内能够因受到盐度的刺激作用而发生促进生长的效果。但当盐度过高或过低时,鱼体往往需要消耗更多的能量用于调整渗透压平衡,而用于生长方面的能量就会减少,当水体盐度处于鱼类生存的极限值附近时,则会出现停止生长或负增长的现象;如果鱼体长期处于这种状态,极易造成其抵抗力下降,并引起死亡。这一结论已在菊黄东方鲀[22]、鲻[23]、日本黄姑鱼[24]等鱼类研究中得到证实。
本研究中,台湾泥鳅幼鱼经过60 d的养殖,生长性能发生了显著性变化(P<0.05)。幼鱼在盐度9‰以上的水体中生活,其特定生长率表现为负值,说明该盐度不适宜其生长。而生活在盐度低于6‰的水体中,能够获得较好的生长率,说明该盐度范围对幼鱼的生长是有利的,此时幼鱼用于生长的能量消耗多于用于维持和调整渗透压平衡所消耗的能量。其中,盐度3‰组幼鱼的各方面指标均最高,提示该盐度是幼鱼的最适生长盐度。
3.3 盐度对台湾泥鳅幼鱼耗氧率的影响
鱼类的生长发育、呼吸代谢等生理活动随外部环境因子的变化而改变,主要包括水温、溶解氧、盐碱度等,其中,盐度对鱼类的新陈代谢有着明显的影响[25-26]。盐度变化首先会引起鱼类的应激反应,紧接着需要调动机体的相关功能进行渗透压调节以平衡体内低渗与体外高渗之间的关系,鱼体在这一过程中需要消耗一定能量并以代谢变化的形式表现出来[27]。鱼类的代谢与盐度的关系可分为:(1)在较大盐度范围内鱼体代谢率相对稳定;(2)机体处于等渗点时代谢率最小;(3)呈“U”形规律,代谢水平在最适盐度两侧都增加;(4)呈“∩”形,代谢率在最适盐度两侧均减小[28]。如浅色黄姑鱼[29]、鲻[30]、褐菖鲉[31]、鱼[32]等在一定盐度范围内,耗氧率随盐度升高逐步上升,在达到峰值后开始下降,表现出了“∩”形变化规律。
本研究中,幼鱼耗氧率在盐度0~6‰范围内逐渐上升,并在盐度为6‰时取得最大值,表明其在该盐度范围内呼吸速率较高,结合生长试验的研究结果(幼鱼在盐度0~6‰范围内生长率较高),推测幼鱼用于自身代谢所消耗的能量可能仅占机体能量消耗的一小部分,而更多能量被用于生长代谢的消耗。其中,鹽度3‰组耗氧率在1 d时即恢复到对照组水平,说明幼鱼对该盐度较为适应。而盐度9‰、12‰组耗氧率一直保持较低水平,尤其是盐度12‰组,这可能是由于水体盐度与幼鱼原来栖息的水体盐度相差太大,该盐度大大超过了它所能适应的盐度范围,为此幼鱼需要消耗更多的能量来平衡和调节渗透压,进而导致幼鱼用于其他方面的能量消耗减少,抑制其渗透压调节功能[33]。
3.4 盐度对台湾泥鳅幼鱼排氨率的影响
当水体盐度发生改变时,会导致鱼类用于调整渗透压平衡的能量增加、新陈代谢加快,从而使耗能增加,进而导致机体排氨率增加[34]。莫荷罗非鱼幼鱼[35]、大黄鱼[26]幼鱼、岱衢族大黄鱼[36]等的排氨率,随着水体盐度的增加表现出先升至最大值后逐渐降低、之后趋于平稳的变化趋势,所不同的是由于试验鱼的种类、规格等不同,排氨率峰值出现时对应的盐度值有所差异。本研究结果与以上结论具有相似性,盐度3‰组幼鱼的排氨率显著高于对照组和其他各盐度组,表明幼鱼在该盐度水体中代谢活动较强,幼鱼由淡水进入低盐环境时能够通过代谢消耗更多的能量以适应外界环境的变化;而对较高盐度则不能适应,比如盐度12‰组排氨率水平一直显著低于其他各试验组的现象则可能说明更高盐度对幼鱼的排泄产生了抑制作用,这一结论与李加儿等对浅色黄姑鱼的研究结论[29]相一致。
本研究所获得的泥鳅幼鱼对低盐和较高盐度的不同适应规律,对盐碱水体的泥鳅养殖生产具有一定的指导意义;同时,泥鳅对不同盐度耐受的生理机制以及盐碱水体中长期养殖效果有待进一步探索和评价。
参考文献:
[1]丁辰龙,王信海,王宣朋,等. 台湾泥鳅与本地泥鳅繁育、性腺发育及生长的比较[J]. 天津师范大学学报(自然科学版),2016,36 (3):69-73.
[2]戴璐怡. 大鳞副泥鳅和台湾泥鳅的形态学研究和线粒体基因组全序列分析[D]. 舟山:浙江海洋学院,2015.
[3]陈石娟. 台湾泥鳅[J]. 海洋与渔业,2014 (1):60.
[4]周 勇,刘桂明,田丽粉. 台湾泥鳅养殖实例[J]. 海洋与渔业,2014 (8):67.
[5]梁德进,黄 维,林碧海. 台湾泥鳅的人工繁殖及育苗技术[J]. 海洋与渔业,2014(7):52-54.
[6]邱楚雯,王韩信,陈迪虎,等. 台湾泥鳅人工繁殖及早期发育的研究[J]. 水产科技情报,2014,41(6):284-289.
[7]尤宏争,邵 蓬,高建忠,等. 出口级黄板鳅(♀)×台湾泥鳅(♂)杂交种及其亲本肌肉营养成分分析比较[J]. 大连海洋大学学报,2017,32(2):198-204.
[8]王 茵,郭书悦,何国森,等. 台湾泥鳅的营养成分分析及评价[J]. 渔业研究,2017,39(5):371-378.
[9]蒲宗旺,王永明,张运邦,等. 台湾泥鳅含肉率及肌肉营养成分分析与评价[J]. 食品工业科技,2017,38(18):300-305,311.
[10]Chiang T Y,Lin H D,Shao K T,et al. Multiple factors have shaped the phylogeography of Chinese spiny loach Cobitis sinensis,in Taiwan as inferred from mitochondrial DNA variation[J]. Journal of Fish Biology,2010,76(5):1173-1189.
[11]刘 静. 盐度对鲤鱼繁殖及相关指标的影响[D]. 大连:大连海洋大学,2015.
[12]杨 建,徐 伟,耿龙武,等. NaHCO3碱度对5种幼鱼的生存及鳃、肾组织的影响[J]. 江西农业大学学报,2014,36(5):1115-1121.
[13]郑伟刚,张兆琪,张美昭. 澎泽鲫幼鱼对盐度和碱度耐受性的研究[J]. 集美大学学报(自然科学版),2004,9(2):127-130.
[14]李洪涛,周文宗,高红莉,等. 运用均匀设计法检验盐度和碱度对泥鳅的联合毒性作用[J]. 水产科学,2006,25(11):563-566.
[15]冯广朋,章龙珍,庄 平,等. 海水网箱养殖长鳍篮子鱼的摄食与生长特性[J]. 海洋渔业,2008,30(1):37-42.
[16]顾 兵,张 政,李玉萍,等. 半数致死量及其计算方法概述[J]. 中国职业医学,2009,36(6):507-508,511.
[17]武鹏飞,耿龙武,姜海峰,等. 三种鳅科鱼对NaCl盐度和NaHCO3碱度的耐受能力[J]. 中国水产科学,2017,24(2):248-257.
[18]杨 建,徐 伟,耿龙武,等. NaHCO3碱度对5种幼鱼的生存及鳃、肾组织的影响[J]. 江西农业大学学报,2014,36(5):1115-1121.
[19]郑伟刚,张兆琪,张美昭,等. 盐度与碱度对花鲈幼鱼的毒性研究[J]. 中国生态农业学报,2005,13(3):116-118.
[20]姚 娜,宋 勇,王 帅,等. 盐度和碱度对塔里木河叶尔羌高原鳅毒性的研究[J]. 西南农业学报,2018,31(2):423-428.
[21]池炳杰,梁利群,刘春雷,等. 滩头雅罗鱼幼鱼对NaCl浓度和碱度的适应性分析[J]. 中国水产科学,2011,18(3):689-694.
[22]施永海,张根玉,刘建忠,等. 低盐对菊黄东方鲀幼鱼生长、存活、耗氧、鳃Na+/K+-ATP酶以及肝抗氧化酶的影响[J]. 动物学杂志,2015,50(3):415-425.
[23]于 娜,李加儿,区又君,等. 盐度胁迫对鲻鱼幼鱼鳃丝Na+/K+-ATP酶活力和体含水量的影响[J]. 动物学杂志,2011,46(1):93-99.
[24]王跃斌,胡则辉,朱云海,等. 盐度对日本黄姑鱼生长及非特异性免疫的影响[J]. 浙江海洋学院学报(自然科学版),2015,34(1):26-31.
[25]Konstantinov A S,Martynova V V. Effect of salinity fluctuations on energetics of juvenile fish[J]. Ichthyol.,1993,33:161-166.
[26]张 涛,王庚申,平洪领,等. 盐度对大黄鱼幼鱼耗氧率和排氨率的影响研究[J]. 浙江海洋学院学报(自然科学版),2015,34(4):310-313.
[27]王兴强,马 甡,董双林. 虾蟹类生物能量学研究进展[J]. 海洋科学,2005,29(10):65-69.
[28]Norman Y S W,Scott P K. Effects of salinity and nutritional status on growth and metabolism of Sparus sarba in a closed seawater system[J]. Aquaculture,1995,135(2):229-238.
[29]李加兒,刘士瑞,区又君,等. 浅色黄姑鱼幼鱼耗氧率、排氨率及窒息点的初步研究[J]. 海洋学报(中文版),2008(5):165-170.
[30]李加儿,曹守花,区又君,等. 温度、盐度和pH对鲻幼鱼耗氧率、排氨率以及窒息点的影响[J]. 中国水产科学,2014,21(5):954-962.
[31]严银龙,袁新程,施永海,等. 盐度对褐菖鲉幼鱼生长、耗氧率和排氨率的影响[J]. 大连海洋大学学报,2019(4):545-550.
[32]闫茂仓,单乐州,邵鑫斌,等. 温度及体重对鱼幼鱼耗氧率和排氨率的影响[J]. 热带海洋学报,2007,26(1):44-49.
[33]闫茂仓,单乐州,谢起浪,等. 温度、盐度及体重对条石鲷幼鱼耗氧率和排氨率的影响[J]. 海洋科学进展,2009,26(4):486-496.
[34]吴庆元. 低盐胁迫对鲻鱼(Mugil cephalus)幼鱼生理生化的影响[D]. 上海:上海海洋大学,2014.
[35]李明昊,陈 刚,黄建盛,等. 盐度对莫荷罗非鱼幼鱼呼吸和氨氮排泄的影响[J]. 广东海洋大学学报,2014,34(1):21-29.
[36]郭念岗,吴雄飞,沈伟良. 温度和盐度对岱衢族大黄鱼排氨氯和耗氧率的影响[J]. 渔业现代化,2014(3):21-25.
3277501908279
猜你喜欢
淡水渔业(2023年2期)2023-03-29 04:03:30
江苏农业科学(2020年17期)2020-10-26 06:54:35
园林科技(2020年2期)2020-01-18 03:28:18
河北渔业(2019年7期)2019-08-27 04:01:48
水产科学(2018年3期)2018-06-04 02:10:58
天津科技大学学报(2016年1期)2016-02-28 16:59:44
广东海洋大学学报(2015年3期)2015-12-22 10:05:30
中国医学科学院学报(2015年5期)2015-03-01 04:03:37
中国医药科学(2015年15期)2015-02-27 12:32:28
应用海洋学学报(2014年1期)2014-11-22 07:17:50
