
胡萝卜叶色突变体叶色参数和色素含量及其相关性分析
2022-03-11 21:10赵宇璇刘真真常双锋徐慧武喆
江苏农业科学 2022年4期
赵宇璇 刘真真 常双锋 徐慧 武喆


摘要:为了探讨胡萝卜不同叶色突变体色差和色素含量的差异及色差与色素含量间的相关关系,以甲基磺酸乙酯(EMS)诱变构建的7个胡萝卜不同叶色突变体为材料,利用色差仪和分光光度计测定和分析突变体叶片的叶色参数(L*、a*、b*)和叶绿素、类胡萝卜素、花色素苷含量。结果表明,胡萝卜不同叶色突变体的叶色参数中亮度值L*与绿叶野生型差异均显著;黄绿叶、紫绿叶、紫红叶和紫叶突变体的色相值a*均为正值,而其余叶色突变体均为负值;黄叶、黄绿叶和浅绿叶突变体的色相值b*均显著高于绿叶野生型,紫叶、紫绿叶突变体均显著低于绿叶野生型。黄绿叶、浅绿叶、深绿叶和紫绿叶突变体总叶绿素的含量极显著高于黄叶和紫叶突变体,深绿叶突变体的类胡萝卜素含量极显著高于其他叶色突变体,紫叶突变体的花色素苷含量最高,极显著高于其他叶色突变体。叶色参数与叶绿素、类胡萝卜素和花色素苷含量的相关性分析结果表明,浅绿叶和深绿叶突变体的叶绿素含量与色相值b*呈极显著相关,黄叶突变体的叶绿素含量与色相值b*呈显著相关,紫绿叶、紫红叶和紫叶突变体的花色素苷含量与色相值a*呈极显著正相关。胡萝卜叶片突变体的色相值a*与肉质根的色相值a*和b*分别呈极显著和显著正相关。
关键词:胡萝卜;突变体;叶色参数;色素含量;相关性分析
中图分类号:S631.201文献标志码: A
文章编号:1002-1302(2022)04-0100-05
收稿日期:2021-04-29
基金项目:山西省重点研发计划(编号:201903D221063);国家自然科学基金(编号:31601751);山西省重点研发计划重点项目(编号:201703D211001-04-01);山西农业大学科技创新基金(编号:2016ZZ02);山西省高等学校科技创新项目(编号:2019L0378)。
作者简介:赵宇璇(1997—),女,山西吕梁人,硕士研究生,主要从事胡萝卜遗传育种与生物技术研究。E-mail:zyx18235448402@163.com。
通信作者:武 喆,博士,副教授,主要从事胡萝卜遗传育种与生物技术研究。E-mail:wzz0618@163.com。
叶色是叶片中不同色素种类和含量综合作用表现出来的结果[1]。叶色突变通常被认为是叶绿素缺陷突变,在自然界中发生频率较高且人工诱变也很容易获得,是辨识度很高的突变类型[2]。一些研究认为叶色突变体致使光合能力下降,植株长势减弱[2]。由于叶绿素含量降低通常使植物的产量和品质下降,过去认为叶色突变对植物生产没有实际意义,往往被剔除掉。但叶绿素是植物进行光合作用的主要色素,主要储存于叶绿体中,因此是研究植物光合作用机制[3]、叶绿素代谢[4]、叶绿体结构和功能[5]的理想材料。同时,叶色突变体也可作为杂交种纯度鉴定的标记性状[6],在辣椒[6]、水稻[7-8]、玉米[9]、甜瓜[10]、菜豆[11]、茄子[12]、黄瓜[13]、羽衣甘蓝[14]等植物上进行了叶色突变体研究。
本试验以甲基磺酸乙酯(EMS)诱变构建的胡萝卜突变体库中的叶色突变体(主要包括黄叶突变体、黄绿叶突变体、浅绿叶突变体、深绿叶突变体、紫绿叶突变体、紫红叶突变体、紫叶突变体)为材料,对突变体的色差(L*、a*、b*)及叶绿素、类胡萝卜素和花色素苷含量进行比较,分析这些色素含量和叶色及根色的相关关系,以期为胡萝卜叶色变化的分子机制研究提供基础。
1 材料与方法
1.1 试验材料
胡萝卜黄叶突变体、黄绿叶突变体、浅绿叶突变体、深绿叶突变体、紫绿叶突变体、紫红叶突变体、紫叶突变体均来源于EMS诱变胡萝卜自交系“17005”构建的胡萝卜突变体库,突变性状表现稳定。
1.2 试验方法
试验时间为2019年3—9月,试验地点位于山西农业大学园艺站(112.55°E,37.68°N)。
试验方法:对胡萝卜野生型及7个突变体品系的叶片色素含量和色差进行测定。
叶片色度的测定参照黄可等的方法[15]。使用日本产Konica Minolta CR-10型色差仪,在光源D65、观察角10°条件下测定不同突变体的叶色参数,每个突变体选取长势、颜色相近的叶片,每张叶片重复测定3次,记录亮度值(L*)、色相值(a*、b*)。其中,L*的取值范围为0~100,L*取值由小到大表明由暗到明;a*表示叶片的红绿程度,取值范围为-128~127,a*值增大,叶片红色加深、绿色减弱;b*表示叶片的黄蓝变化,取值范围为-128~127,叶片的b*值增加,叶片蓝色减弱,黄色增强[16]。
叶绿素、类胡萝卜素含量的测定和计算方法参照李合生的方法[17],每个样品重复3次;花色素苷含量的测定和计算参照Pirie等的方法[18-19]。
1.3 数据分析
通过Excel 2016进行数据处理,采用SPSS 23.0软件进行方差分析,用最小显著性差异(LSD)法进行多重比较,用Origin 2019b作图软件作图。
2 结果与分析
2.1 不同叶色突变体的叶色变异
由图1可知,野生型的叶片为绿色(图1-a),突变体叶片的颜色有黄叶(图1-b)、黄绿叶(图1-c)、浅绿叶(图1-d)、深绿叶(图1-e)、紫绿叶(图1-f)、紫红叶(图1-g)和紫叶(圖1-h)。叶片颜色突变有全叶突变和部分突变。
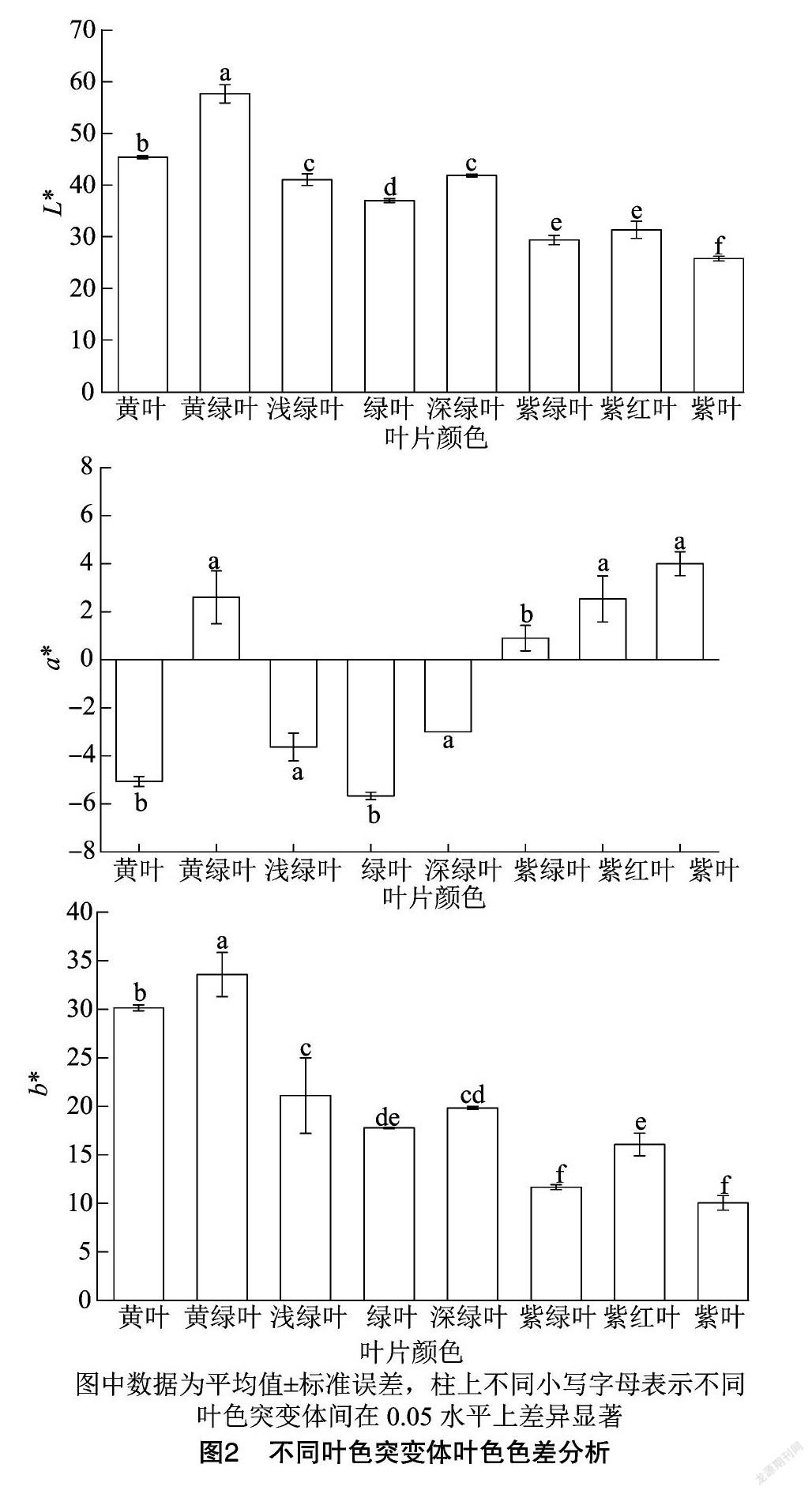
2.2 不同叶色突变体叶片色差分析
叶色参数是利用色差计来定量表征色泽在三维空间中变化的变量值,7个叶色突变体的相关指标如图2所示。在7个叶色突变体中,叶片亮度变化规律为黄绿叶突变体的叶片亮度最大,黄叶突变体次之,而紫叶突变体的亮度最低,不同叶色突变体的亮度值(L*)与绿叶野生型差异均显著(P<0.05)。色差指标a*表示从绿到红的变化,不同叶色突变体呈现不同的a*值,黄绿叶、紫红叶和紫叶突变体的a*值较为接近,差异不显著,浅绿叶和深绿叶的a*值与绿叶野生型的a*值差异达到显著水平(P<0.05)。色差指标b*表示从蓝到黄的变化,其中,黄绿叶突变体的b*值最大,约为33,黄叶突变体次之,紫叶突变体的b*值最小,仅为11,除深绿叶突变体外,其他叶色突变体的b*值与绿叶野生型的差异均达到显著水平(P<0.05)。
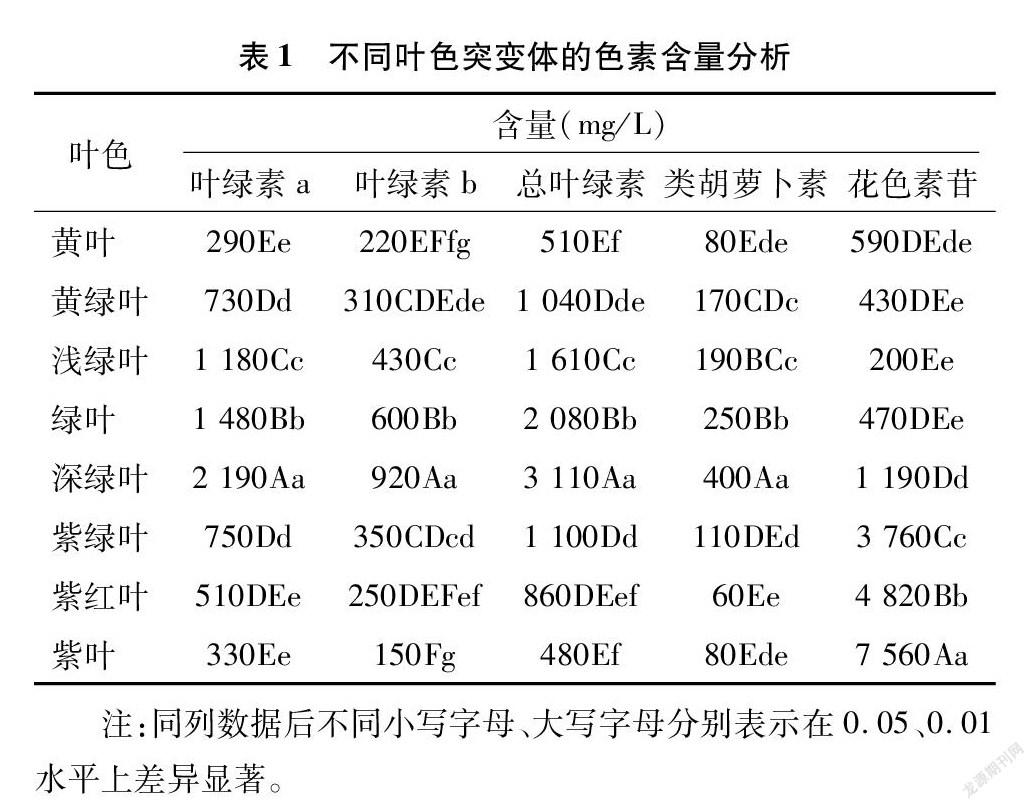
2.3 不同叶色突变体色素含量分析
由表1可知,深绿叶突变体的叶绿素a含量最高,黄叶突变体含量最少,且与绿叶野生型差异达极显著水平(P<0.01)。深绿叶突变体的叶绿素b含量也极显著高于绿叶野生型,其余叶色突变体的叶绿素b含量都极显著低于绿叶野生型(P<0.01)。深绿叶突变体的总叶绿素含量和类胡萝卜素含量最高,分别达到 311、 400 mg/L, 极显著高于绿叶野生型。黄叶、紫红叶和紫叶突变体的总叶绿素含量均极显著低于绿叶野生型,但三者之间差异不显著,黄绿叶、浅绿叶和紫绿叶的含量居中,但也都极显著低于绿叶野生型。紫红叶突变体的类胡萝卜素含量最低,黄叶和紫叶突变体次之,三者之间差异不显著,黄绿叶和紫绿叶突变体的类胡萝卜素含量均极显著低于绿叶野生型(P<0.01),浅绿叶突变体与绿叶野生型之间差异显著(P<0.05)。紫绿叶、紫红叶和紫叶突变体中花色素苷的含量极显著高于其他突变体和野生型,其中紫叶突变体最高,紫红叶次之,紫绿叶最低,三者之间差异达到极显著水平(P<0.01)。
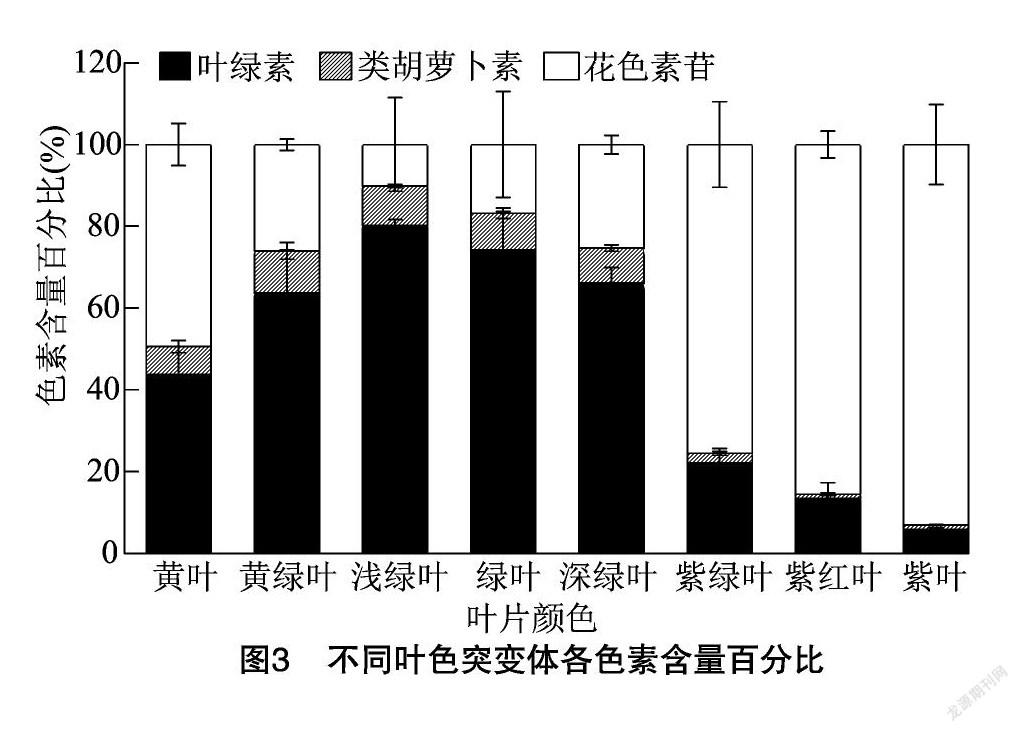
2.4 不同叶色突变体各种色素含量的百分比
由图3可以看出,绿叶野生型、黄绿叶、浅绿叶和深绿叶突变体中叶绿素含量的百分比都达到60%以上,而紫绿叶、紫红叶和紫叶突变体中花色素苷的比例占60%以上,黄叶突变体中叶绿素和花色素苷含量的占比都在40%以上,类胡萝卜素含量在所有叶片类型中的占比都较低。
2.5 色素含量与叶色参数的相关性分析
由表2可以看出,浅绿叶突变体的L*值与叶绿素含量呈极显著相关,深绿叶和紫绿叶突变体的L*值与类胡萝卜素和叶绿素含量都呈显著或极显著相关;浅绿叶突变体的a*值与叶绿素含量呈极显著负相关,绿叶野生型的a*值与花色素苷含量呈显著负相关,深绿叶突变体的a*值与类胡萝卜素和叶绿素含量均呈极显著负相关,紫绿叶、紫红叶和紫叶突变体的a*值均与花色素苷含量呈极显著正相关,紫绿叶突变体的a*值还与类胡萝卜素含量呈显著正相关,紫叶突变体的a*值与类胡萝卜素和叶绿素含量呈显著或极显著正相关;黄叶和浅绿叶突变体的b*值都与叶绿素含量呈显著或极显著负相关,绿叶野生型的b*值与类胡萝卜素、叶绿素和花色素苷含量呈极显著负相关,深绿叶的b*值与类胡萝卜素和叶绿素呈极显著正相关,紫红叶和紫叶的b*值与类胡萝卜素呈显著负相关。黄叶、黄绿叶和浅绿叶突变体的3个叶色参数与类胡萝卜素含量均没有显著相关性,因此类胡萝卜素含量并不影响这3个突变体的叶色,但类胡萝卜素含量对其他叶色突变类型的叶色均有不同程度的影响。
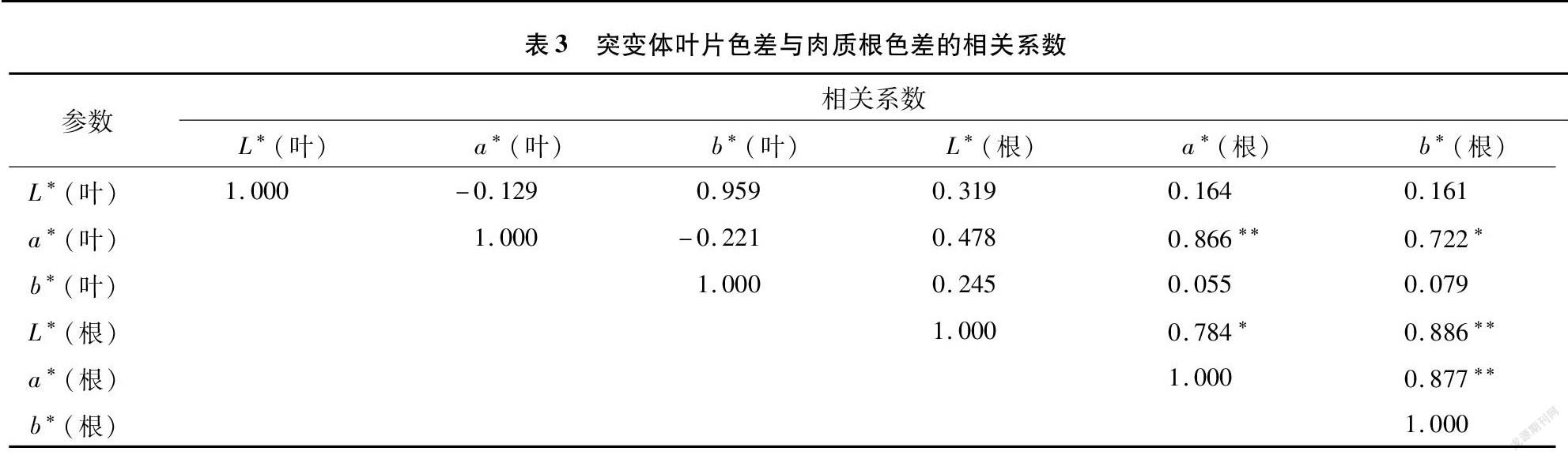
2.6 叶片色差与肉质根色差的相关关系
由表3可知,胡萝卜突变体叶片的a*值与肉质根的a*值呈极显著正相关,与肉质根的b*值呈显著正相关。肉质根的L*值与肉质根的a*值和b*值呈显著或极显著正相关,肉质根的a*值与b*值之间的相关性也达到极显著水平。这说明胡萝卜地上部叶片色差参数a*值对地下部肉质根的a*值和b*值有一定的影响,而叶色参数L*值并不是影响肉质根叶色参数的主要因素。
3 讨论与结论
3.1 不同叶色突变体叶色参数及色素含量变化
利用色差仪测定的叶色参数已广泛应用于植物叶片叶色光泽度的量化分析,克服了靠感官定性描述叶色的缺陷[20]。本研究中不同叶色突变体的L*值均为正值,与绿叶野生型的差异均显著;不同叶色突变体呈现不同的a*值,黄绿叶、紫红叶和紫叶突变体的a*值差异不显著,浅绿叶和深绿叶的a*值与绿叶野生型的差异达到显著水平;黄绿叶突变体的b*值最大,黄叶突变体次之,紫叶突变体的b*值最小,除深绿叶和紫红叶突变体外,其他叶色突变体的b*值与绿叶野生型的差异均达到显著水平。
叶色是叶片中不同色素种类和含量综合作用表现出的结果[2]。叶绿素、类胡萝卜素和花色素苷等色素都属于次生代谢物质,它们的比例决定着植物叶片器官的着色[21]。本试验中,不同叶色突变体的叶绿素、类胡萝卜素和花色素苷3种色素的比例不同,因此叶片呈现的颜色也不同。7种叶色突变体中,紫叶突变体的花色素苷含量占比最大,所以叶片呈现紫色,而黄绿叶、浅绿叶、深绿叶突变体和绿叶野生型的叶绿素含量比例高于花色素苷含量,因此叶片展现绿色,黄叶突变体的叶绿素和花色素苷含量占比相近。 李云飞解剖彩叶植物組织发现,花色素苷在植物中分布不同会影响叶片呈色[22],本试验结果与之一致。
3.2 不同叶色突变体叶色参数与色素含量的相关关系
许多植物的叶色参数与色素种类及含量之间存在显著相关关系[23],但不同类型植物色素变化不同,叶色参数也存在差异[24]。本研究中紫色叶片类型的突变体花色素苷含量较高,叶色参数的a*值与其呈极显著正相关,说明花色素苷含量影响这种突变体的a*值。浅绿叶和深绿叶突变体的L*值、a*值和b*值均与叶绿素含量呈极显著相关,说明叶绿素含量影响这2个突变体叶色的L*值、a*值和b*值。黄叶、黄绿叶和浅绿叶突变体的3个叶色参数与类胡萝卜素含量均没有显著相关性,因此类胡萝卜素并不影响这3个突变体的叶色。朱书香等对4种李属彩叶植物色素含量与叶色参数的相关性研究表明,色素变化趋势基本相同,但色素含量和叶色参数的相关性却因树种各异而不同[23],本试验结果与之相似。
参考文献:
[1]赵昶灵,郭华春.植物花色苷生物合成酶类的亚细胞组织研究进展[J]. 西北植物学报,2007,27(8):1695-1701.
[2]徐明远,何 鹏,赖 伟,等. 植物叶色变异分子机制研究进展[J]. 分子植物育种,2021,19(10):3448-3455.
[3]Fambrini M, Castagna A,Dalla Vecchia F,et al. Characterization of a pigment-deficient mutant of sunflower(Helianthus annuus L.) with abnormal chloroplast biogenesis,reduced PSⅡ activity and low endogenous level of abscisic acid[J]. Plant Science,2004,167(1):79-89.
[4]Parks B M,Quail P H.Phytochrome-deficient hy1 and hy2 long hypocotyl mutants of Arabidopsis are defective in phytochrome chromophore biosynthesis[J]. The Plant Cell,1991,3(11):1177-1186.
[5]Agrawal G K,Yamazaki M,Kobayashi M,et al. Screening of the rice viviparous mutants generated by endogenous retrotransposon Tos17 insertion.Tagging of a zeaxanthin epoxidase gene and a novel OsTATC gene[J]. Plant Physiology,2001,125(3):1248-1257.
[6]马志虎,颜素芳,胡志中,等. 辣椒黄绿苗突变体生物学特性及生长动态研究[J]. 种子,2000(4):10-12.
[7]叶 俊,吴建国,杜 婧,等. 水稻“9311”突变体筛选和突变体库构建[J]. 作物学报,2006,32(10):1525-1529,1599-1600.
[8]蔡星星,张 盛,王 欢,等. 水稻株型基因的研究现状及应用前景[J]. 分子植物育种,2017,15(7):2809-2814.
[9]赵 琦,唐崇钦,匡廷云.玉米突变体(zb/zb)的叶绿体光合特性[J]. 植物学报,1997,39(11):1082-1084.
[10]宋燕妮,杜黎黎,王学征,等. 甜瓜突变体库的构建及M2群体表型变异的研究[J]. 植物遗传资源学报,2015,16(6):1338-1344.
[11]陈其福,刘 畅,斯琴图雅,等. 菜豆A18-160Co-γ射线诱变和突变体库的构建[J]. 中国蔬菜,2019(5):38-44.
[12]刘富中,张 映,杨锦坤,等. 茄子叶色黄化突变体的特征及遗传分析[J]. 园艺学报,2020,47(12):2340-2348.
[13]国艳梅,顾兴芳,张春震,等. 黄瓜叶色突变体遗传机制的研究[J]. 园艺学报,2003,30(4):409-412.
[14]陈安和,李加纳,柴友荣,等. 羽衣甘蓝中一个突变型肉桂酸-4-羟化酶基因的克隆及分析[J]. 园艺学报,2007,34(4):915-922.
[15]黄 可,王小德,柳翼飞,等. 红枫春季叶色变化与色素含量的相关性[J]. 浙江农林大学学报,2012,29(5):734-738.
[16]McCallum C M,Comai L,Greene E A,et al. Targeted screening for induced mutations[J]. Nature Biotechnology,2000,18(4):455-457.
[17]李合生.植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:135-137.
[18]Pirie A,Mullins M G.Changes in anthocyanin and phenolics content of grapevine leaf and fruit tissues treated with sucrose,nitrate,and abscisic acid[J]. Plant Physiology,1976,58(4):468-472.
[19]Xu J R,Zhang M W,Liu X H,et al. Correlation between antioxidation and the content of total phenolics and anthocyanin in black soybean accessions[J]. Agricultural Sciences in China,2007,6(2):150-158.
[20]董建文,廖艷梅,许贤书,等. 秋季观赏植物单株美景度评价[J]. 东北林业大学学报,2010,38(3):42-46.
[21]李雪飞,韩甜甜,董 彦,等. 紫叶稠李叶片色素及氮含量与其光谱反射特性的相关性[J]. 林业科学,2011,47(8):75-81.
[22]李云飞. 土壤干旱胁迫对李属彩叶植物抗旱生理及叶色的影响[D]. 保定:河北农业大学,2008.
[23]朱书香,杨建民,王中华,等. 4种李属彩叶植物色素含量与叶色参数的关系[J]. 西北植物学报,2009,29(8):1663-1669.
[24]葛雨萱,王亮生,周肖红,等. 香山黄栌叶色和色素组成的相互关系及时空变化[J]. 林业科学,2011,47(4):38-42.
3283500338220
猜你喜欢
今日农业(2021年7期)2021-11-27
今日农业(2021年15期)2021-11-26
今日农业(2020年15期)2020-12-15
安徽医科大学学报(2016年12期)2017-01-15
时代金融(2016年29期)2016-12-05
山东农业工程学院学报(2016年6期)2016-12-01
中国市场(2016年40期)2016-11-28
商(2016年33期)2016-11-24
天津医科大学学报(2015年2期)2015-12-22
山东医药(2015年40期)2015-02-28
