
紫花苜蓿粗灰分与矿质元素含量QTL定位分析
2022-03-08 01:05:54杨习江杨昌福蒋学乾杨青川康俊梅
草地学报 2022年2期
杨习江, 杨昌福, 蒋学乾, 张 帆, 何 飞, 杨青川, 康俊梅
(中国农业科学院北京畜牧兽医研究所, 北京 100089)
植物体内钾(Potassium,K)、钙(Calcium,Ca)、镁(Magnesium,Mg)、磷(Phosphorus,P)等矿质元素是促进植物生长发育的重要营养元素[1],也是牧草矿质营养的重要组成部分。牧草作为草食动物的重要食物来源,其体内的矿质元素对动物的营养调节代谢[2]发挥重要的作用。K是植物中最丰富的阳离子,参与植物渗透压、细胞膜电位、pH的调节过程[3],也参与蛋白质合成和光合作用过程中酶的活化[4],同时也能参与动物的葡萄糖转运[5]。有研究发现,施K可提高紫花苜蓿(Medicagosativa)的碳水化合物含量,并调控其在苜蓿各器官中的合理分配,进而增强苜蓿对蓟马(Thripidae)的耐害性[6]。Ca是生物体中普遍存在的第二信使,它参与各种细胞过程,调控植物的发育,以及响应生物和非生物胁迫响应[7],比如对植物干旱、伤害和热胁迫的快速反应,从而增强植物的免疫力[8]。施加适宜浓度的CaCl2可以缓解干旱对苜蓿幼苗造成的伤害[9]。Ca也是动物骨骼和牙齿的主要成分。Mg是叶绿素的中心元素,是一系列固碳相关酶的辅助因子。因此,Mg与植物光合作用和光合产物的运输密切相关,是植物生长发育的重要元素[10]。在动物中,Mg对肌肉再生也有很重要的作用[11]。P与植物产量调控密切相关,有研究报道当植物对P的摄入不足时会使植株产量降低10%~15%[12],适宜施P量可有效提高苜蓿种子荚果数、结荚率和种子产量[13]。动物体内P是必不可少的营养元素,是细胞膜、三磷酸腺苷(ATP)、骨骼和牙齿的主要构成成份,主要参与能量代谢,脂类物质的吸收和转运,以及动物蛋白的形成等生物学功能。粗灰分含量是衡量饲料产品和饲料原料中营养成分的重要指标之一,也是饲料产品必须测定的营养指标之一,粗灰分测定对指导饲料及饲料原料的加工都具有重要意义[14]。因此,研究牧草体内矿物质元素的遗传信息不仅对揭示其自身的生物学功能奠定理论基础,而且对通过牧草的遗传改良为草食动物提供营养元素具有重要意义。
植物体内矿物质元素的传统测定方法为化学测定法,该方法耗时长、成本昂贵。近红外光谱技术(Near infrared spectrum instrument,NIRS)具有无损、快速等优点,近年来已被广泛应用于植物的品质相关性状的测定[15-17]。由于植物的大多数品质相关性状,以及矿质元素(如K,Ca,Mg,P等)含量都是由多个基因调控的数量性状,既受环境的影响,也受环境和基因型互作的影响[18]。针对数量性状位点的研究,利用连锁分析方法能够有效地鉴定出控制这些复杂性状的数量性状基因座(Quantitative trait loci,QTL)。到目前为止,在很多植物中开展了关于矿质元素的QTL研究,如水稻(Oryzasativa)[19-20],玉米(Zeamays)[21-22],小麦(Triticumaestivum)[23-24],大豆(Glycinemax)[25]和花生(Arachishypogaea)[26]。
紫花苜蓿是世界上种植面积最大的多年生豆科牧草,由于产量高、品质优、营养价值高,具有丰富的矿物质元素等特点,被誉为“牧草之王”[27-28]。苜蓿的品质相关性状一直是育种家关注的研究重点,国内外对于苜蓿品质相关性状已经开展了大量的研究,主要集中在蛋白质、次生代谢物、纤维素、木质素等方面[18,29-30],与矿质元素相关的研究鲜有报道,只有一些初步的关联分析[18]和基因组预测[30]。本研究以紫花苜蓿F1杂交群体为材料,采用近红外光谱技术分析法对苜蓿的粗灰分及矿质元素含量进行测定,结合原有的高密度遗传图谱,初步检测与粗灰分,K,Ca,Mg,P品质性状相关的QTL位点,找到一些稳定遗传的分子标记,以期为挖掘调控苜蓿矿质元素相关候选基因奠定基础,同时紧密连锁的分子标记可用于分子标记辅助选择(Marker-assisted selection,MAS)育种对苜蓿进行遗传改良。
1 材料与方法
1.1 试验地概况
试验地位于河北省廊坊市中国农业科学院(万庄) 国际农业高新技术产业园基地(39.59°N,116.59°E),属于暖温带大陆性季风气候,四季分明,冬寒夏热,秋凉春旱。年均气温11.9℃左右,年均降水量554.9 mm,年均日照时数2 660 h左右,无霜期183 d。供试土壤质地为中性土壤,有机质含量为1.69%,pH值为7.37。
1.2 试验材料
本试验以低产早熟紫花苜蓿为父本(P1,CF000735),以高产晚熟紫花苜蓿为母本(P2,CF032020),且在矿质元素含量上有显著差异的父母本进行种内杂交,创建了包含392个单株的F1作图群体。F1群体种子于2015年在温室培养。通过扦插获得亲本和子代的无性繁殖株系,在2016年早春,将双亲和F1群体移栽到田间种植,设置3个重复,行距1 m,株距0.8 m。扦插后生长至初花期前刈割两次,确保植株的生长一致性,之后再进行植株表型的测定。
1.3 表型性状测定与统计分析
分别于2016,2019和2020年的初花期刈割第一茬植株。初花期即单株观察到第一朵小花的开放时期,时间大概在每年5月,不同单株刈割时间不同。然后将样品置于鼓风干燥箱内,60℃条件下烘24 h至绝干。用粉碎机研磨,过1 mm筛网后,取150 g粉样进行标号装袋。随后,采用近红外反射光谱仪(FOSNIRS1650)对苜蓿中的粗灰分(Ash),K,Ca,Mg和P含量进行测定。2017和2018年由于人员不足,无法完成大规模粉碎及测定工作,因此未收集到表型数据。在测定前,对仪器模型进行校正,用10%的样品进行化学法测定,与近红外光谱仪器测定数据进行比较和校准,误差在10%以内。测定数据用Microsoft Excel 2019软件进行整理,用SAS 9.4软件进行基本统计分析和相关性分析。
1.4 遗传图谱与QTL定位分析
利用本实验室前期已构建好的紫花苜蓿的遗传连锁图谱[31]为基础,进行粗灰分与4种矿质元素的QTL定位。构建好的父本连锁图谱包含32个连锁群,944个单核苷酸多态性(Single nucleotide polymorphism,SNP)标记,图谱总长4 088.70 cM,标记间的平均遗传距离4.33 cM。母本连锁图谱包含32个连锁群, 2 874个SNP标记,图谱总长4 229.15 cM,标记间的平均遗传距离1.50 cM。使用QTL IciMapping软件,首先进行遗传力和最佳线性无偏差预测值(Best linear unbiased prediction,BLUP)的计算;之后利用BIP功能,选择ICIM-ADD模型,以LOD > 3为阈值,进行QTL检测(QTL命名遵循McCouch等[32]原则);最后用Mapchart 2.3软件绘制QTL定位图。
2 结果与分析
2.1 粗灰分与矿质元素表型分析
对F1群体及亲本在3年测定的粗灰分与4种矿质元素性状表型值进行统计分析(表1),发现除去2016年的Ash,K和P含量,以及2020年P含量父母本之间未表现出显著性差异,在其他环境下父本性状均优于母本,且存在显著性差异(P<0.05)。F1群体中,粗灰分与4种矿质元素K,Ca,Mg,P含量分别占干物质含量的7.0%,2.0%,1.5%,0.3%和0.3%,且从变异幅度看,其分布范围均在双亲差异的范围之外,存在广泛变异(表1)。同时,粗灰分与4种矿质元素含量性状的遗传力均大于0.50,具有较高的遗传力,表明受遗传特性的影响较大。另外,除2019年测定的Ash,K,Mg含量峰度和偏度绝对值偏差较大外,其余性状在群体中分布的峰度和偏度绝对值均小于1(表1),其表型频率分布均表现为正态或近似正态的连续分布,且存在双向超亲分离现象,符合QTL定位分析。
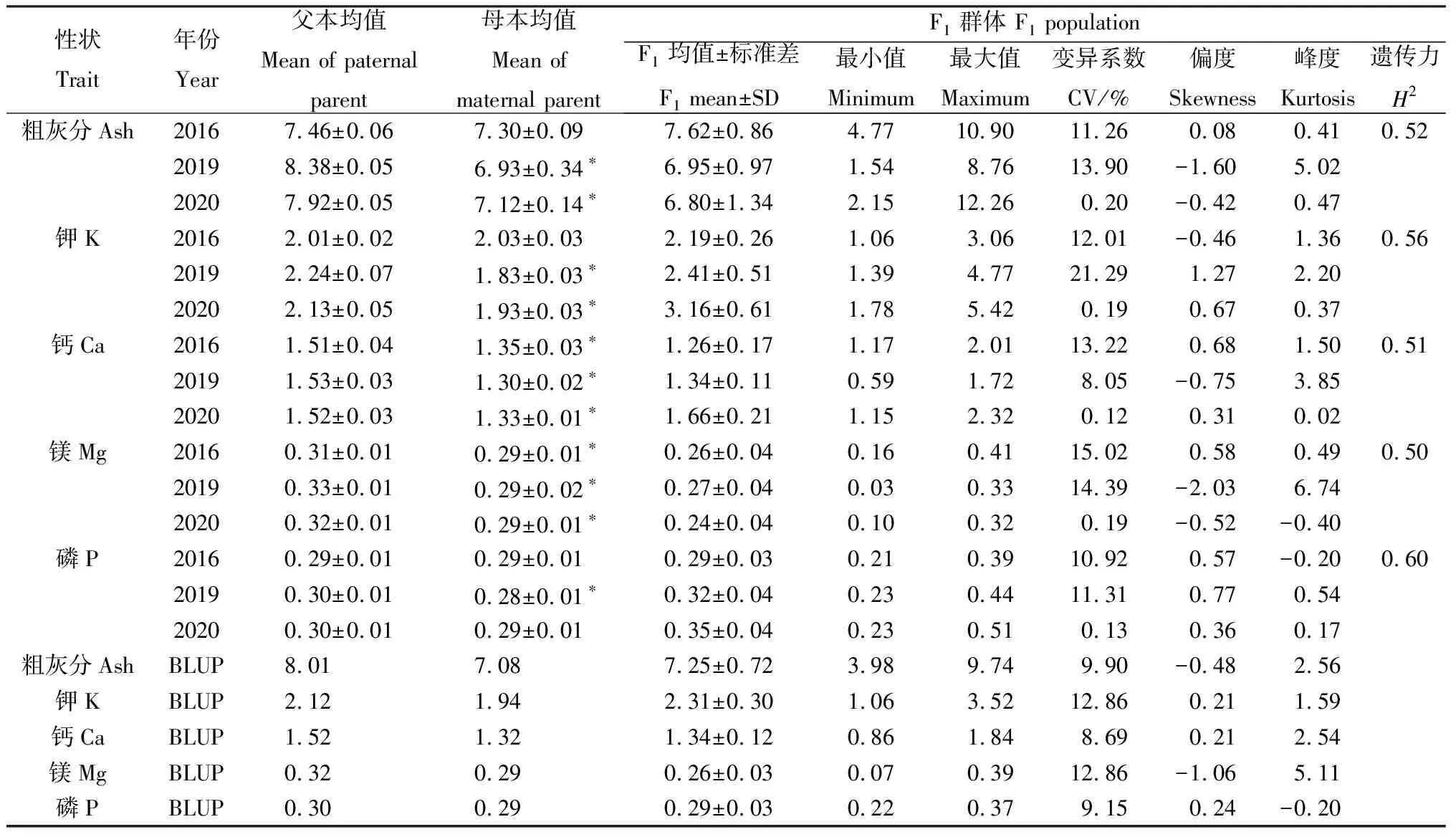
表1 粗灰分和4种矿质元素在F1群体中的统计Table 1 Statistics of ash and four mineral elements in F1population
2.2 粗灰分与矿质元素含量间的相关性分析
对F1群体粗灰分与4种矿质元素含量间相关性进行分析(表2),发现除Mg含量和Ca含量相关性较低外(P<0.05),其余性状间都呈极显著正或负相关关系(P<0.001)(表2)。Ash含量与Mg含量极显著正相关(P<0.001)(r=0.608),却与K,Ca,P含量极显著负相关(P<0.001)。矿质元素之间,除Mg含量与K和Ca含量呈负相关关系外(P<0.05),其余K含量与Ca和P含量呈极显著正相关关系(P<0.001),P含量与Ca和Mg含量也呈极显著正相关关系(P<0.001)。

表2 F1群体5个品质相关性状间的Pearson相关性分析Table 2 Pearson correlation analysis for five quality-related traits in F1 populations
2.3 QTL定位分析
基于前期构建的高密度遗传图谱,3个环境中共检测出63个与粗灰分和4种矿质元素,含量相关QTL(表3,图1,图2),分布于2C,3C,4B,4D,6B,6D,7D,8B染色体的不同位置,贡献率介于3.04%~24.83%之间。经鉴定与粗灰分相关的QTL共10个,其中3个位于父本,7个位于母本,贡献率介于2.88%~5.10%之间。母本中qAsh-8B-1和qAsh-8B-2属于重复定位QTL,位于8B染色体125 cM处,侧翼标记区间为TP65937-TP6047,贡献率分别为5.06%和4.71%。3个环境中共鉴定出10个与K含量相关的QTL,均来源于母本连锁群,解释的表型变异为3.10%~7.25%,其中于2020年环境中检测到表型变异最大的QTL(qK-2C),位于2C染色体TP77362-TP23382标记区间内,可解释7.25%的表型变异。共检测到15个与Ca含量相关的QTL,3个位于父本,12个位于母本,贡献率介于3.00%~6.56%之间。重复定位的QTL共有6个:其中qCa-3C-1和qCa-3C-2位于父本3C染色体TP29222-TP47945标记区间内,分别可解释3.95%和3.04%的表型变异;qCa-6B-1和qCa-6B-2位于母本6B染色体TP2660-TP96620标记区间内,贡献率分别为3.00%和3.72%;qCa-6D-1和qCa-6D-2位于母本6D染色体TP8104-TP24250标记区间内,分别可解释6.14%和3.64%的表型变异。所有重复定位QTL的加性效应值均为负(Add < 0),表明影响Ca含量增加的位点均来自于父本。三个环境下与Mg相关的QTL共12个,可解释表型变异为3.42%~12.77%。其中有3个QTL的贡献率大于10%,分别是qMg-6D-2,qMg-7D,qMg-8B,贡献率分别为12.77%,10.00%,10.97%。此外,16个与P含量相关QTL被鉴定,贡献率介于2.50%~29.85%。其中有7个QTL贡献率大于10%,其中qP-6D-1,qP-6D-2,qP-6D-3三个位于母本6D染色体,贡献率分别为21.56%,29.85%,11.12%;qP-7D-2,qP-7D-3,qP-7D-4三个位于父本7D染色体,分别可解释11.84%,20.77%,24.83%的表型变异;qP-4D位于父本4D染色体,贡献率为10.11%。
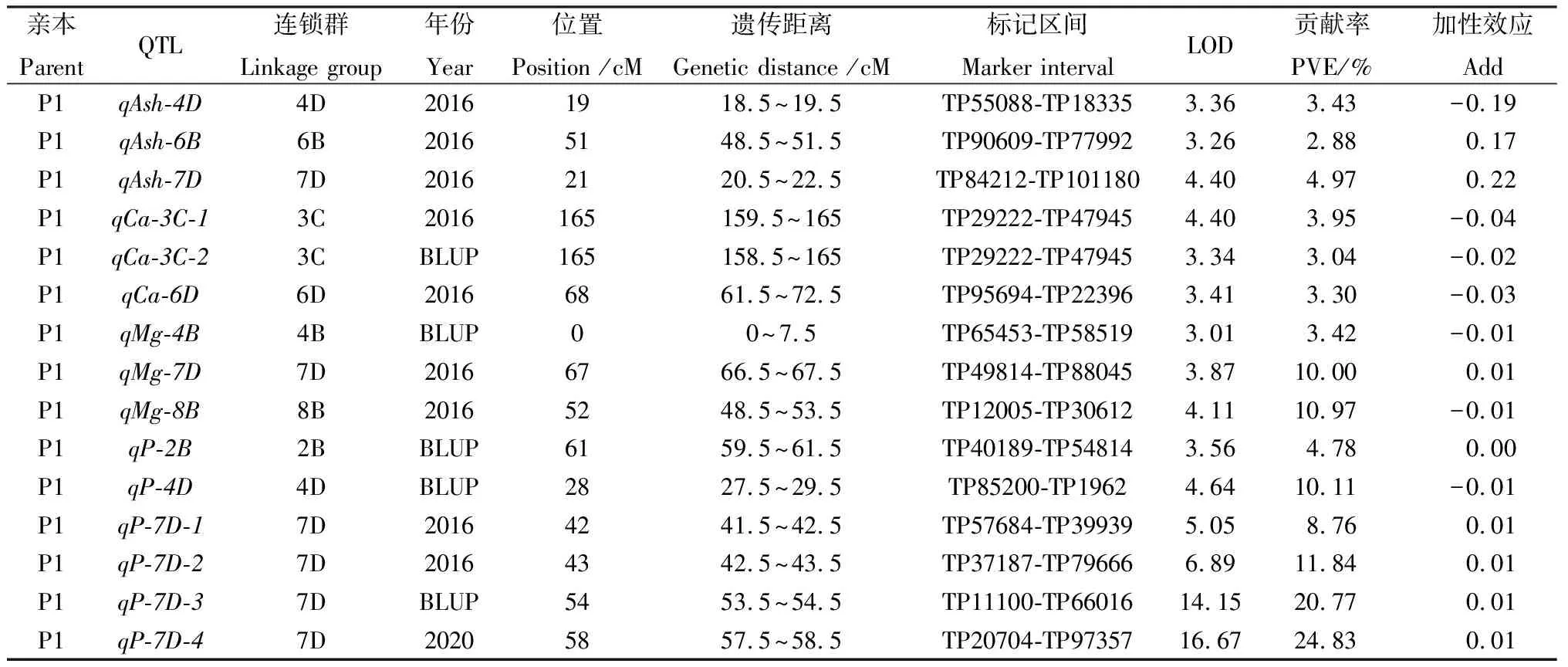
表3 紫花苜蓿F1群体五个品质性状的QTL定位结果Table 3 QTL analysis results of five quality traits in F1 population of alfalfa
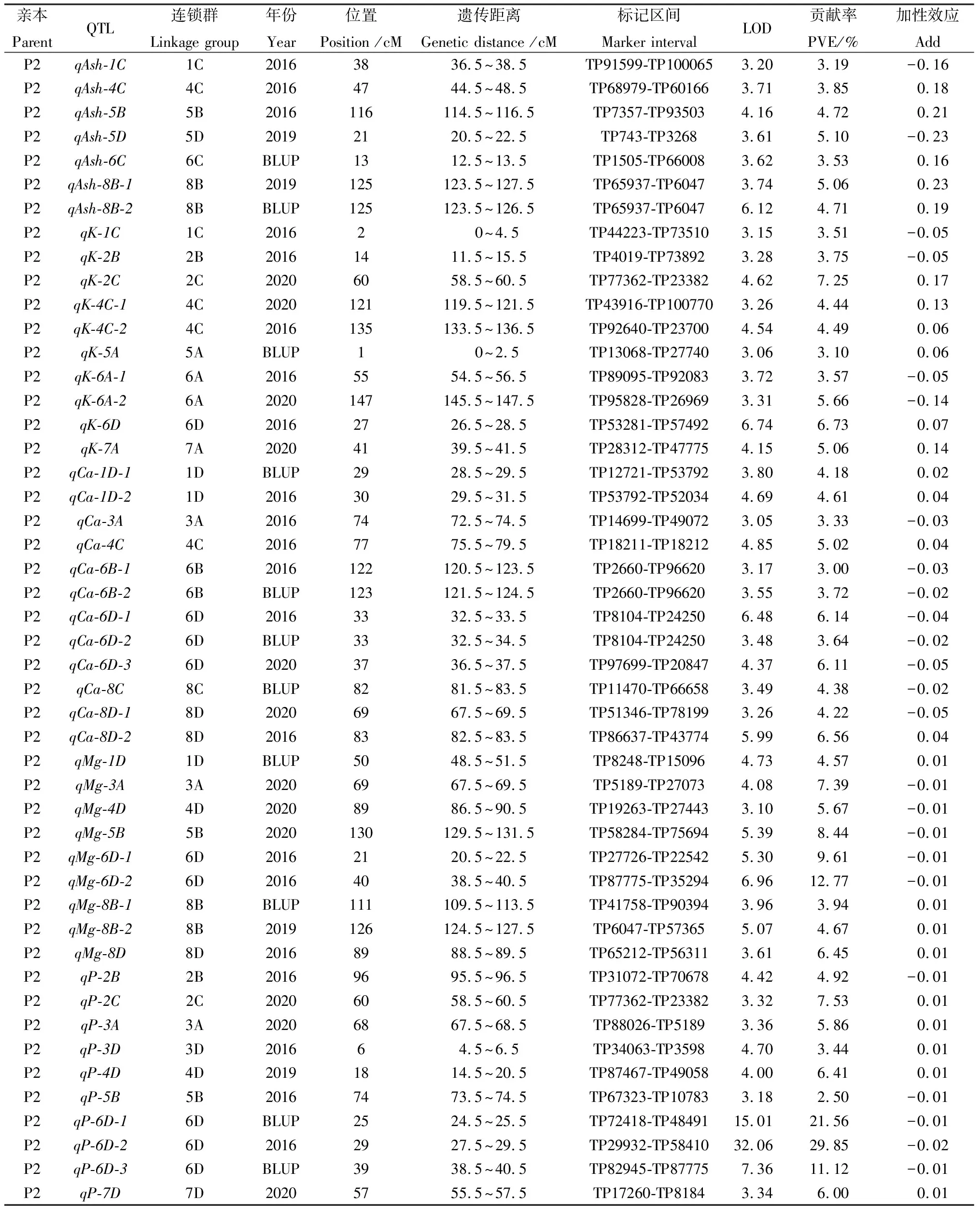
续表3
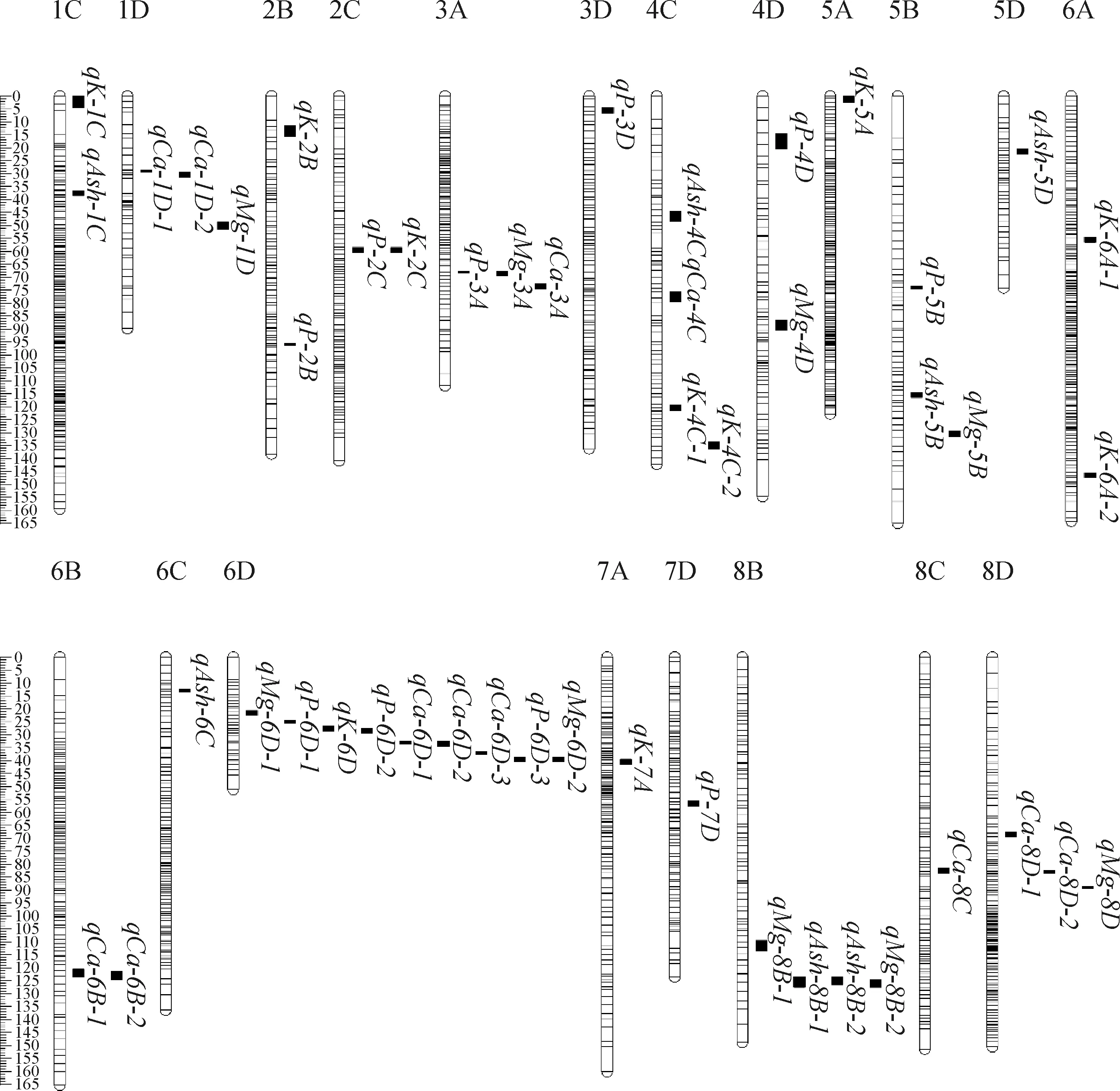
图2 苜蓿5种品质性状QTL在母本遗传连锁群上的分布情况Fig.2 Distribution of QTLs for five quality traits on genetic map of maternal parent in alfalfa
此外,鉴定了3个共定位区间,其中qP-2C和qK-2C共定位于母本2C染色体60 cM处,分别解释7.53%和7.25%的表型变异;qP-3A和qMg-3A共定位于母本3A染色体上67.5~69.5cM区间,贡献率分别为5.86%和7.39%;qP-6D-3和qMg-6D-2共定位于母本6D染色体上38.5~40.5 cM区间,贡献率分别为11.12%和12.77%。
3 讨论
对苜蓿粗灰分与矿质元素含量测定数据分析发现,粗灰分与4种矿质元素K,Ca,Mg,P含量分别占干物质含量的7%,2%,1.5%,0.3%和0.3%,与以往关于苜蓿品质相关性状含量测定的结果一致[18]。考虑到各个性状受到一因多效性或多个基因相互作用的影响[33],对粗灰分及4种矿质元素进行了相关性分析。结果显示,粗灰分含量与K,Ca,Mg,P含量均存在显著的相关性,这与矿质元素是粗灰分的主要组分有直接关系。此外,研究发现Mg含量与K,Ca含量都呈显著负相关关系,K含量却与Ca含量为显著正相关关系,这可能是因为K,Ca,Mg参与了离子转运载体的竞争,3者之间存在拮抗作用所造成的[34-36]。
QTL的多效性指同一区间检测到的QTL同时参与几个性状的调控,这种情况在动植物中都普遍存在[37-38],且此类与多个性状关联的标记在今后的分子标记辅助选择上将会更有效地用于改善植株的多个性状。本研究的结果也证实了多效应QTL的存在。在母本2C连锁群上,qP-2C和qK-2C都定位于60 cM处,分别解释了7.53%和7.25%的表型变异,表明TP77362-TP23382标记区间内可能存在一个QTL与P和K含量均有相关性;在母本3A 和6D染色体上区间67.5~69.5 cM和38.5~40.5 cM处,存在两组与P和Mg含量相关的多效应QTL,分别为qP-3A和qMg-3A,qP-6D-3和qMg-6D-2,解释表型变异较高,这些被定位在相同或相邻区间的QTL与表型性状间的相关性具有高度一致性。
此外,本试验检测到一些成簇分布的QTL,位于母本6D染色体上的成簇QTL有9个,集中分布于21~40 cM区间,其中与K含量相关的有1个,与Ca含量相关的有3个,与Mg含量相关的有2个,与P含量相关的有3个;位于父本7D染色体上的成簇QTL有6个,集中分布于21~67 cM区间,其中与Ash含量相关的有1个,与P含量相关的有4个,与Mg含量相关的有1个。成簇QTL产生的原因可能有两点:一是连锁群本身比其他连锁群长度短,标记密度大(分别为0.50 cM和1.64 cM),所以更容易检测出QTL;二是由于一因多效或微效基因紧密连锁于同一区间或基因重叠[39],使相关性状的QTL通常定位于同一连锁群上相同或者相近区域。这些QTL能在不同环境或遗传背景下,在不同性状被检测到,说明其表达具有一定的稳定、可靠性,将对苜蓿矿质元素相关分子标记研究具有重要意义。随着紫花苜蓿基因组的破译[40-41],本研究中检测出的多效应QTL和成簇分布的QTL区间将在今后研究中得到进一步验证,通过鉴定紧密连锁的分子标记,将为苜蓿分子标记辅助选择育种提供重要的标记。
4 结论
本研究共检测到63个与粗灰分,K,Ca,Mg,P含量相关的QTL,其中在父本7D染色体和母本6D染色体上存在QTL富集区域,分别检测出6个和9个QTL,且贡献率较高,今后如果对这些遗传区域进行深入研究,挖掘与主效QTL紧密连锁的分子标记,可为利用分子标记辅助选择育种培育更丰富矿质元素的苜蓿新品种提供理论指导。
猜你喜欢
选煤技术(2022年2期)2022-06-06 09:13:10
选煤技术(2022年2期)2022-06-06 09:12:22
选煤技术(2021年6期)2021-04-19 12:22:08
四川蚕业(2021年4期)2021-03-08 02:59:56
中成药(2018年11期)2018-11-24 02:57:08
现代园艺(2017年21期)2018-01-03 06:41:41
中国种业(2016年11期)2016-12-01 05:43:51
食品工业科技(2014年9期)2014-03-11 18:15:44
植物营养与肥料学报(2011年3期)2011-10-24 06:14:44
种子科技(2010年5期)2010-05-28 09:24:34
