
基于生物多样性的樟江流域自然保护地空间优化
2022-03-04 12:50傅微甄广润
生态科学 2022年1期
傅微, 甄广润
基于生物多样性的樟江流域自然保护地空间优化
傅微,*, 甄广润
北京建筑大学建筑与城市规划学院, 北京 100044
优化生态保护空间格局是我国生态文明建设的重要内容, 也是国际保护生态学研究的前沿方向。评估资源禀赋、识别并确定优先次序和预测保护区对保护和管理濒危物种至关重要。樟江流域内已占11.9%流域面积的保护区仍出现77.9%流域面积的生境显著退化, 发展与保护亟待合理平衡。Zonation4GUI软件以互补性非穷尽式算法为基础, 在优化已有保护网络并表征稀有性和独特性方面结果较为理想, 以此展开樟江流域自然保护地空间整合优化路径研究。先人工预处理物种栖息地分布格局、物种保护权重、规划单元, 并迭代选择输入性生境退化条件图层、保护代价图层, 结合人工干扰情况提出三种不同情景下的优化方案。结果表明: 保护优先区对代理物种栖息地有良好的覆盖效果, 生态完整性优先情景和最小社会经济和土地资源代价的保护优化情景均呈现扩建小于8%的一级优先区域, 所有保护对象均受到保护; 加入27%左右的所有优先区域, 两种优化方案覆盖所有保护对象栖息地的比例提升70.0%和59.0%。虽然生态完整性优先的优化情景优于最小社会经济和土地资源代价下的优化情景, 但保护代价更高。人为干扰强烈的抢救优先的优化情景中, 90.7%的保护对象栖息地覆盖范围有明显提升。从技术和操作层面, 为重构中国自然保护地体系整合优化方案提供路径和方法。
系统保护规划; 保护空缺; 喀斯特地区; 流域整体保护; 保护目标
0 前言
生物多样性为人类的福祉和长期生存提供生态系统服务和商品, 保持生态系统稳定性, 维持生态平衡[1-2], 是全球生态系统的不可分割的要素。生物多样性在农业、林业、渔业和旅游业等推动经济发展中发挥着重要作用。人口和人均消费的持续增长导致了不可持续的开发利用生物多样性, 物种的过度开发、外来入侵物种的扩张、气候变化、森林退化和独特栖息地的破坏等驱动力严重影响生物多样性[3]。随着生物多样性丧失危机的不断加剧, 保护生物多样性的意识和政策机制正在区域、国家和国际层面增强[4], 旨在最小化物种退化和生境破坏。为了增加保护工作的可行性, 并考虑到世界上大多数地区的生物多样性清单都不完整, 通常使用物种多样性代理确定支持多个物种和栖息地的保护区系统保护规划[5-6]。在过去的三十年里, 许多研究采用多样性格局代理(如物种丰富度)作为最小覆盖集合表征研究区内几乎所有的物种[7], 选址潜在的自然保护区。根据物种丰富度选择保护地是最传统的方法之一(案例可见[8-9])。然而, 当保护规划的目标旨在优化已有保护网络并表征关键区域的最大物种数量时, 基于物种丰富度并非保护优先级筛选的良好指标, 因其无法反映互补性(Complementarity), 即一种基于场地的保护价值衡量方法, 确保被选择纳入保护区网络的地点补充已有保护空间; 同时无法说明特有性和稀缺性, 从而出现受保护物种的数量减少现象[10]。互补算法可解释物种稀有性, 选择的地点是相互补充的, 每个地点不一定是物种最丰富的, 但每个地点在物种组成上都不同, 可产生一种包含所有物种的地点的解决方案, 在有限的范围内给予所有物种更高的优先权[11]。
樟江流域呈扇形展布, 地跨贵州省黔南布依族苗族自治州荔波、三都县, 属于珠江流域。主河道樟江发源于贵州省荔波县佳荣乡月亮山, 由东北向西南流, 流域面积1673.9 km2。樟江流域是中国生物多样性热点地区之一, 景观类型发育具有典型性、稀有性、脆弱性以及多样性, 表现在世界范围内喀斯特地貌上保存完好的区域, 是长江、珠江上游重要的生态屏障。其丰富的、独特的生物多样性为流域上游、下游提供生态系统服务和商品, 但由于其脆弱性和资源密集性, 易受到包括过度开发在内的各种驱动力变化因素的影响。虽然已建立了5个自然保护地, 但这些保护地提供的保护并不总是有效的[12]。土地开发和环境变化导致栖息地生境减少, 可能影响物种的分布, 经济发展和生物多样性保护的竞争需求也存在着激烈矛盾冲突[13]。
在此背景下, 以樟江流域为研究区域旨在关注并回答以下问题: 近20年已有自然保护地的保护效益是否足够?人类开发利用的干扰程度及与之带来的生境退化趋势如何?采用互补算法最大化保证濒危物种数量和最优化补充保护空间; 筛选考虑不同保护策略的保护空缺格局和保护效果。
1 整合优化路径与方法
1.1 自然禀赋研究
1.1.1 对象选择
依据外业调查、专家知识, 选取研究区内重点保护对象54种, 其中植物38种, 动物16种, 如表1所示。植物物种相关信息及分布来自中国科学院植物研究所中国珍稀濒危植物信息系统(http:// www.iplant.cn/rep/), 动物物种相关信息及分布来源自中国动物主题库(http://www.zoology.csdb.cn/), 并于2017年9月至2018年1月和2018年12月开展外业工作予以核实。

表1 重点保护物种列表
1.1.2 空间分布
空间分布采用统计物种分布模型(Statistical Species Distribution Models), 该模型结合了物种名录、自然生态系统与资源环境数据, 提供栖息地适宜性的连续图层预测[14]。首先查找物种分布范围, 再依据栖息地类型对分布范围进行修正: 植物物种的栖息地类型通过海拔高程、土壤类型、植物类型制图叠加生成; 动物物种栖息地类型则通过植被类型、海拔高程及土地利用类型制图叠加获取, 并在ArcGIS支持下叠加生成物种适宜空间分布图。此外, 提取每种保护对象的适宜栖息地面积, 作为保护目标分析指标。植被类型分布来源于《1:1 000 000 中国植被图集》矢量化数据; 土壤类型空间分布数据来源于中国科学院资源环境科学数据中心(http:// www.resdc.cn); 数字高程模型SRTM_DEM数据(30 m分辨率), 来源于http://srtm.csi.cgiar.org/。
1.1.3 保护资源权重
依据自然保护地中典型性、稀有性、自然性、多样性、科学价值、美学价值(观赏价值)、社会和经济价值等指标, 整合保护对象权重。如公式(1)所示, 选用国家保护等级、地方特有性、物种濒危程度、经济价值和潜在栖息地面积五个指数制定权重方案。
=(endangered+protection+endemic+economic+area)/5 (1)
式(1)中是物种的保护权重;endangered按物种的濒危程度打分;protection按国家保护等级,endemic特有性打分;economic按照植物的经济用途打分, 很多植物具有药用、色素、野菜、观赏、油脂、纤维、蜜源、香料、野果等经济价值。area按物种潜在栖息地面积打分, 最高分5分。动物不考虑经济价值, 去掉economic后四项取平均值[15]。
1.1.4 规划单元
为突出流域生态系统结构的完整性以及流域上下游之间的连续性, 规划单元采用ArcGIS Hydrology工具构建1421个集水区, 因为集水区单元具有更大的自然相似性, 符合物种的自然分布特征, 可维持保护对象生境和地貌单元的完整性。
1.2 保护现状梳理
流域内已有樟江国家级重点风景名胜区、国家级茂兰喀斯特森林自然保护区和3个县级自然保护区(含核心区、缓冲区及实验区), 分布数据来源于黔南州林业局根据实地调查记录经纬度信息在ArcGIS10.2中空间成图, 作为去除掩膜图层, 为完善保护优化格局提供基础。
1.2.1 流域自然保护现状条件
流域自然保护现状条件采用植被净初级生产力2000年–2015年年平均值趋势变化作为指标表征生境特征退化情况。植被净初级生产力为生态系统在特定时期内的生物总量, 表征生态质量, 是评价生态系统功能特征的重要参量。数据基于CASA (Carnegie Ames Stanford Approach)模型计算, 采用逐像元一元线性回归分析方法, 阐释2000—2015年研究区植被净初级生产力的时间序列变化趋势, 基于斜率和Pearson相关系数判定退化程度。此外, 结合《樟江流域综合规划》资料, 评估河流物理特征、水文水资源特征、水质状况、水生生物、社会服务功能情况, 绘制流域自然保护现状景观条件图。生境退化条件情况中条件值可以在0和1之间变化, 值为1表示原始状态, 生境适宜性或特征发生没有退化; 0的值表示完全退化的状态, 任何负值(丢失的数据)都将被视为零。
1.2.2 人为干扰度量与保护代价成本
基于生物多样性的保护优化格局完整性的实现受制于人类活动空间的干扰, 为维护完整性而改变人为干扰空间所需保护代价成本随人为影响程度增加而增加。因此参照相关研究[16], 选取公路、铁路、城镇、农村居民点、水坝作为计算规划单元保护代价的因子, 将每个因子的度量标准化后, 赋予不同的权重(表2)。相加计算得到每个规划单元的保护代价指数。将保护代价指数用保护区覆盖面积比例进行调整, 得到最终保护代价成本分布图(cost layer)。道路、居民点数据、土地利用数据均来源于黔南州国土部门2017年土地利用数据; 大坝分布来源于《樟江流域综合规划》, 计算如公式(2-3)所示。

表2 各影响因子的度量和权重
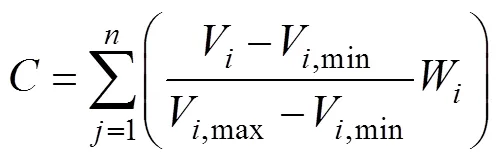
C′=C(1–0.5) (3)
式(2)中:V为每个保护单元内因子的度量值;W为因子的权重;为每个保护单元内因子的个数; C为每个保护单元调整前的代价值; 式(3)中C′为每个保护单元经过保护区调整后的代价值;为每个保护单元内被保护区覆盖的面积比。
1.3 结构优化分析
1.3.1 保护价值评估
保护价值评估基于空间保护优先级划分和大尺度空间保护规划的前沿软件Zonation4GUI, 基于互补的算法对单元进行分层排序[11]。首先假设所有单元都可能是重要的, 并在每次迭代后逐步删除最不重要的单元。通过最小化保护价值的边际损失, 最大程度降低生物多样性损失, 并考虑到保护对象及分布的典型性、稀有性、多样性、高价值等特征的连通性需求和优先级。最终保证整体连通性为焦点找出最能配合现有保护地的范围, 即最有保护价值的地区[17]。
1.3.2 分析保护空缺
本研究区域原有自然保护地(Protected Area, 简称PA)占总面积的11.9%, 因此, 选取樟江流域景观总面积的30%作为目标优先区, 将最高保护价值区域的10%划分为一级优先区, 10%—20%划分为二级优先区, 20%—30%划分为三级优先区, 最终产生不同级别的保护优先格局。
1.4 优化整合方案
分别将前述优化路径中保护对象空间分布图层、物种名录、保护权重、保护规划单元图层、已有自然保护地图层、自然保护现状条件图层、流域范围图层代入Zonation4GUI软件, 选择性代入保护代价图层作为不同保护策略情景展开分析。采用了保持格局连通性的核心区移除规则(Core Area Zonation, 简称CAZ) 和边缘移除规则(Rule of Edge Removal)[18], 分层优先逐步得到一个高连通性景观结构的嵌套序列。进一步研究保护空缺区域与周边自然保护地的关系, 将生态系统类型相同、主要保护对象和保护目标一致的保护空缺区域补充并入周边自然保护地, 设立新的自然保护地。
分层优化过程中, 引入三种不同保护策略情景下的保护空间优化方案, (1)生态完整性优先的优化方案, 该方案不考虑保护代价迭代, 将高保护价值规划单元建立优先保护格局, 最大程度降低生物多样性损失; (2)最小的社会经济和土地资源代价下优化方案, 该方案考虑保护代价, 并作为成本层参与迭代分析, 平衡利益相关者和建立较为可行的保护区; (3)抢救性保护的优化方案, 选取人为干扰强烈的分布区叠加生态完整性高保护价值区域作为优先保护格局。
2 结果与分析
2.1 流域保护现状堪忧
当现状自然保护区、风景名胜区和世界遗产地等已有自然保护地总面积为199.1 km2的情形下, 樟江流域生态退化现状如图1所示, 生境显著退化面积置信度超过99%的退化面积占流域总面积的46.8%, 置信度超过95%的显著退化面积占流域总面积的67.0%, 置信度超过90%的显著退化面积占总面积的77.9%。可见, 樟江流域近16年时间内生态退化明显且严峻, 水源涵养能力下降, 流域的生态环境质量呈逐年下降趋势, 生态脆弱性加剧。根据樟江荔波水文站的资料, 生态流量干流部分退化25%, 从河流连通性、河道蜿蜒度、河道渠化程度、河岸稳定性、河床稳定性、河流形态指标表明河流物理结构干流退化了31.2%, 上游水源区退化了7.3%, 导致水体污染, 水量减少等现象。
根据适宜栖息地分布格局, 动物栖息地分布面积居高, 其中穿山甲、普通鵟、小鸦鹃、褐翅鸦鹃、小灵猫和金猫分布较广, 辐花苣苔、中国蕨、白花兜兰、单座苣苔、狭叶含笑等分布少, 辐花苣苔分布适宜范围为899 m2。已有保护区内不同类别物种栖息地分布情况发现, 半枫荷、单性木兰、辐花苣苔、福建柏、红豆杉、黄杉、楠木、异性玉叶金花、中国蕨尚未覆盖, 其他79.6%的覆盖物种栖息地面积比例均较低, 平均比例为7.1%, 表明现有保护地空间保护情况有较大不足。
保护代价成本分析结果显示(图2), 樟江流域改变需要的保护代价高的规划单元较集中分布在樟江流域中线地带——荔波县城樟江干流、上游居民点集中区为轴线, 以及分布于三荔水库大坝、的马水库大坝集水区区域。
2.2 生态完整性优先的优化方案
如图3所示, 高保护价值且存在保护空缺的区域集中分布在樟江主河道两岸亚热带常绿阔叶林带、樟江源头区域拉易河、爽姑河汇入水昔河的交汇处、水令河上游、爽姑河上游、拉滩河上游, 茂兰镇瑶麓喀斯特典型生态系统亚热带常绿阔叶林区域。这些地区物种类型分布丰富, 具有较好的优先级。一、二、三级优先区共补充保护空缺面积453.6 km2(表3), 统计每个物种的栖息地在保护优先区内分布情况, 共计45个物种的栖息地面积比例超过60%, 约占全部物种的83.3%, 实现对森林生态环境、生物多样性、珍稀野生动植物和水源涵养林地的保护。其中, 一级优先区范围内新增面积比例为7.4%, 即可实现涵盖100%研究选取保护对象的部分或者全部适宜栖息地, 其中金猫、猕猴、普通鵟等鸟类和哺乳类动物分布空间比例最高, 对如辐花苣苔等此类适宜栖息地面积极小的物种也实现其栖息地的完全保护。二级优先区范围内新增面积比9.7%, 进一步实现了单座苣苔、红豆杉、楠木、四药门花、狭叶含笑、中国蕨保护对象栖息地生境的完全保护。三级优先区范围内不足总面积的0.0033%为原有保护区范围, 继续实现了福建柏、蓝翅八色鸫、单性木花、翠柏、白花兜兰栖息地生境的完全保护。
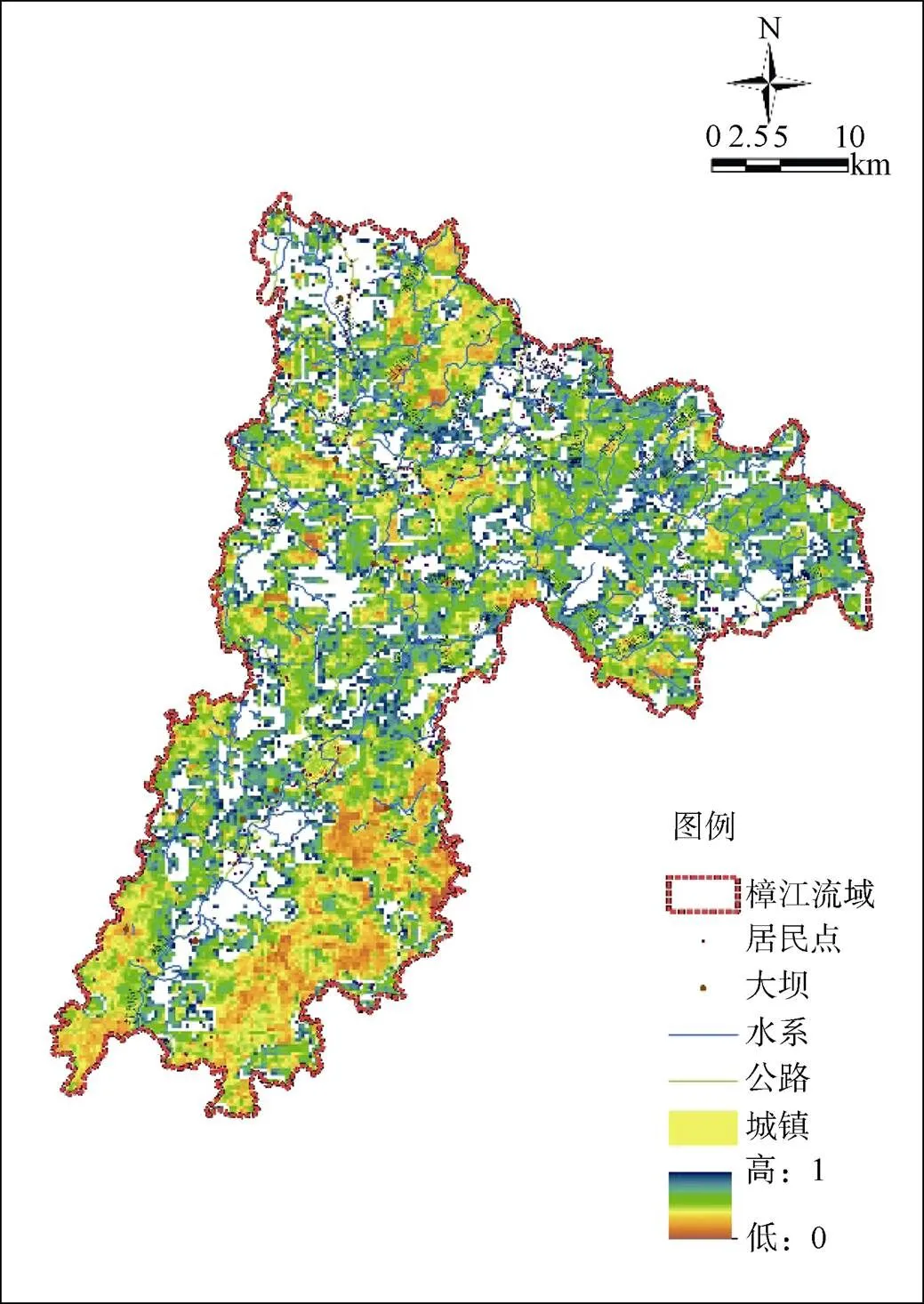
图 1 生态退化显著性分布
Figure 1 Distribution of condition layer

图 2 保护代价成本分布
Figure 2 Distribution of cost layer
2.3 最小社会经济和土地资源代价的优化方案
最小的社会经济和土地资源代价下最大程度开展保护方案, 高保护价值区域集中分布在樟江主河道两岸亚热带常绿阔叶林地带、水便河中下游、樟江源头区域水昔河上游拉滩河集水区、爽姑河汇入水昔河的交汇处以及水令河上游(图4)。通过互补算法Zonation软件分析所得优化空间, 虽然相对于动物的适宜性栖息地分布, 部分植被保护对象表现出可适应的栖息地面积非常有限, 但在纳入一级优先区后, 已覆盖全部38种植物和16种动物, 新增优先区面积占流域总面积的8.0%。植物物种栖息地所占比例最少的4个对象为辐花苣苔、狭叶含笑、红豆杉、单座苣苔、中国蕨, 面积分别为75 m2、257 m2、275 m2、276 m2、1235 m2。补充纳入二级优先区内也同样全覆盖保护对象, 保护空缺占流域总面积9.5%, 辐花苣苔适宜栖息地得以完全保护。补充纳入三级优先区作为辅助补充提高物种可栖息地的面积比例, 保护空缺占比9.7%, 生物多样性代理保护对象中, 单座苣苔、辐花苣苔、福建柏、红豆杉、中国蕨的栖息地完全覆盖。一、二、三级优先区共新增优化空间454.7 km2, 共计32个物种的栖息地面积比例超过50%、22个物种的栖息地面积超过60%, 该保护方案可进一步实现环境保护与经济发展同步开展。

图 3 生态完整性优先保护方案
Figure 3 Landscape rankings for scenarios1
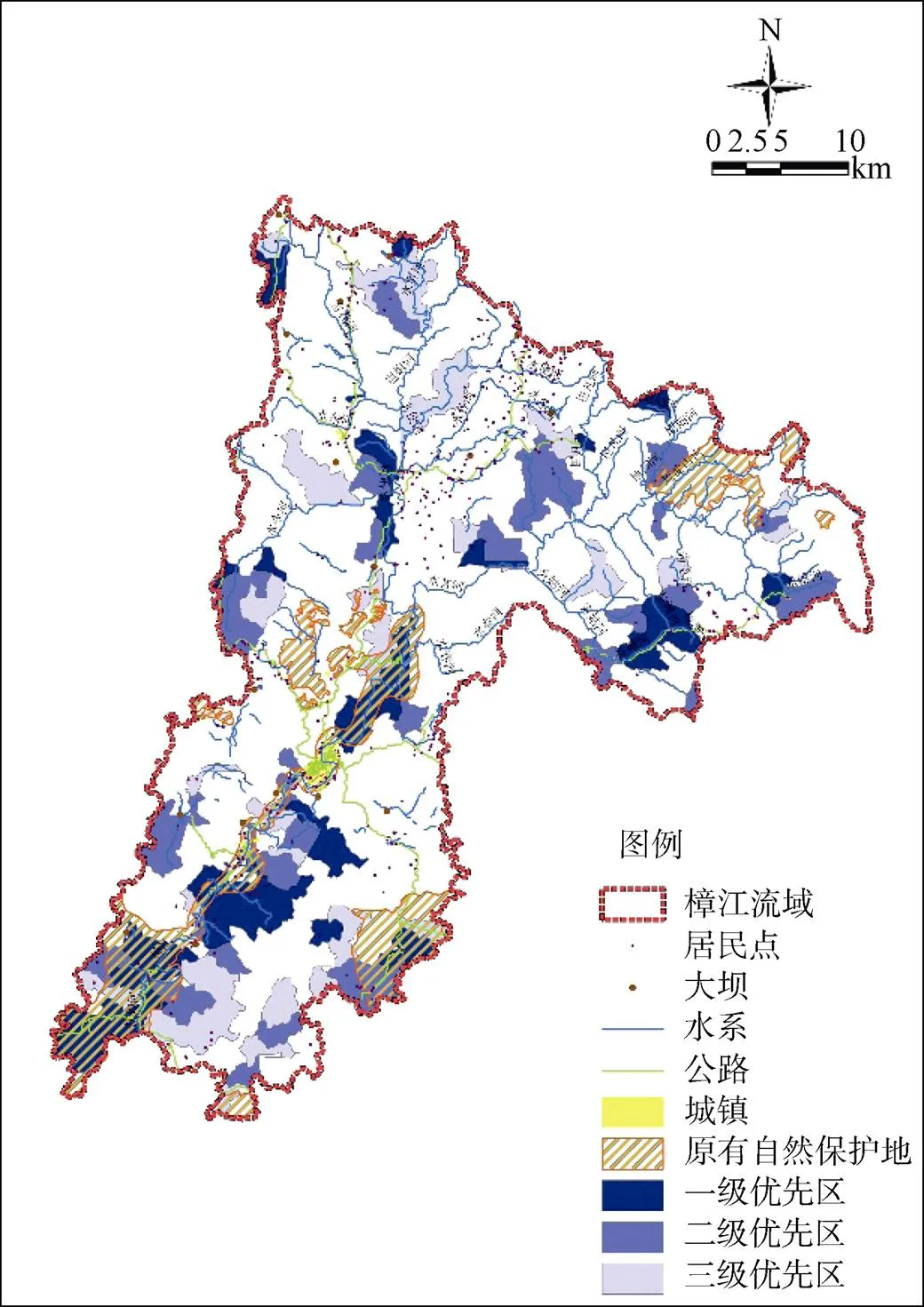
图 4 最小的社会经济和土地资源代价下保护方案
Figure 4 Landscape rankings for scenarios2
2.4 抢救性保护的优化方案
抢救优先的保护方案中(图5和表3), 一、二、三级优先区共新增优化空间126.6 km2, 约占流域总面积的7.6%, 保护的物种栖息地除翠柏、红豆杉、单座苣苔、楠木5类植物外, 均有涉及。空间优先区主要分布于沿樟江主河道两岸地带, 这一水陆交接地带从栖息地分析结果发现存在超过半数代表性动植物物种和特有种, 表明了这一地带存在抢救性保护空缺, 研究结果也与政府报告中提到主河道两岸缓冲区保护效率不高相吻合。此外, 一级优先区中, 保护空缺面积为29.2 km2, 主要分布于水贾河上游集水区; 二级优先区保护空缺面积39.3 km2, 主要分布于水各河和水令河上游集水区; 三级优先区保护空缺面积58.0 km2, 集中于水昔河和水便河上游山谷集水区地带, 也是人为干扰明显区域。
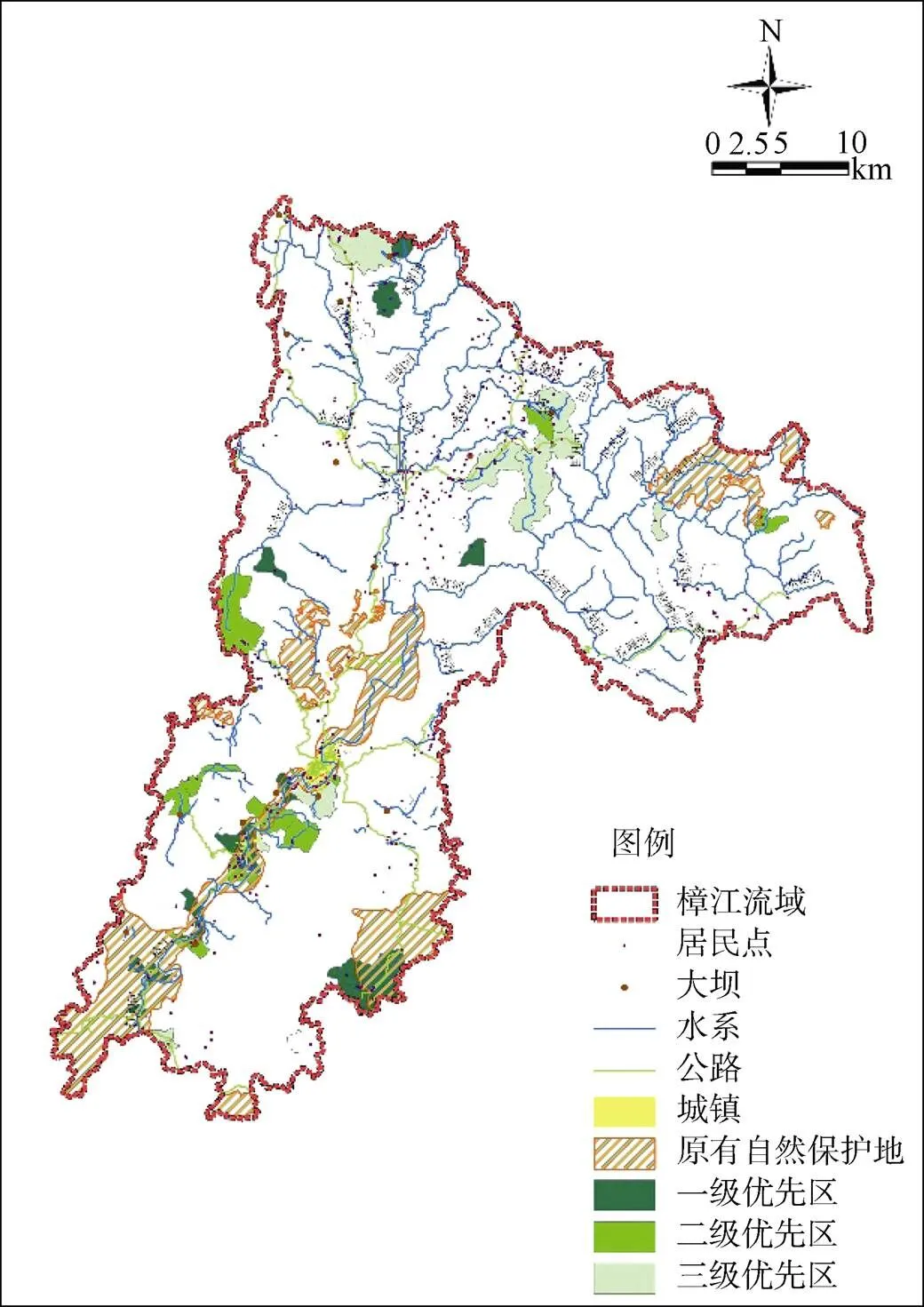
图 5 抢救性优先保护方案
Figure 5 Landscape rankings for scenarios3

表3 三种预景方案下分级保护优先区的分布比例与面积(单位: km2)
3 讨论
研究揭示了至少两个值得探讨的方面: 首先, 用于保护规划的资源通常是有限的, 这就要求采取尽可能高效的保护行动, 以最大限度地保护生物多样性[7]。在确定物种的空间保护优先权及地点保护价值时, 相对于物种丰富度的穷尽式算法, 互补性算法是非穷尽式的, 即并非极致追求物种最丰富的地点, 而是综合物种组成, 集中代表所有的物种[11]。对比近年来采用物种丰富度算法的研究发现, 探究已有保护区与算法模拟保护区展开额外重叠分析[19], 但从实际建设可行性层面, 互补性算法可在已有保护区的范围内优化新增, 而非调整原有保护区微分, 显著提高了景观空间优先化的有效性。互补算法Zonation的核心区域分区(CAZ), 也对物种稀有性和独特性给予了较好的解析, 因为其优先考虑对物种重要的核心区域, 测试了所有可能的地点选择场景, 在有限的范围内给予那些物种更高的优先权。近三十年来, 保护生物学家已经认识到互补性比物种丰富度更有效; 然而, 它尚未成为生物多样性保护研究的主流方法[20-21], Astudillo‑Scalia等[22]学者猜测是因为这个概念还没有被很好地理解, 特别是基于典型区域应用实践层面。
其次, 通过比较保护代价层介入与否的系统保护规划的有效性, 选择作为保护优先级的空间分布有明显差别。取决于人类干扰对生态完整性造成的影响, 未纳入保护代价的保护目标的实现情况在所需保护面积小于纳入后。因此, 人为干扰加剧条件下, 未来所需扩大保护地的范围也有待相应提高。人为干扰强烈的抢救性优先保护区初步建议开展生境恢复试点, 改善其生态功能。缓解自然保护地与社区之间在自然资源利用上的矛盾, 社区共管[23]与生态旅游[24]在这些地带的重要体现, 补充收入来源, 开展环境容量评估。寻找保护与周边用地、产业发展的平衡点, 较大程度改善矛盾冲突的核心地带。确定优先保护区域是生物多样性保护的重要第一步, 但关键是根据这些建议采取行动[25], 樟江流域在自然保护地空间范畴与管理范畴需协调一致的努力。
4 结论
近16年樟江流域生境退化置信度超过95%的面积占流域总面积的67.0%, 流域内生物多样性资源交易频繁, 许多物种栖息地分布逐渐破碎, 特别是水系干流和上游水源涵养地带仍未得到保护。已有保护区覆盖代理保护对象栖息地面积比例不足8%, 基于生物多样性的樟江流域自然保护地空间优化迫在眉睫, 是保护地体系建设、管理和决策的重要依据。采用Zonation核心区域分区(CAZ)优化路径, 对栖息地分布小、物种丰富度不足的物种保护互补性较好。基于三种不同情景下的优化结果表明, 保护优先区对代理物种栖息地有良好的覆盖效果, 生态完整性优先情景和最小社会经济和土地资源代价的保护优化情景均呈现: 在现有格局中加入小于8%的1级优先区域, 所有代理物种均受到保护。加入27%左右的所有优先区域, 两种优化方案覆盖所有保护对象栖息地的比例提升70.0%和59.0%。虽然生态完整性优先的优化情景优于最小社会经济和土地资源代价下的优化情景, 但保护代价更高。人为干扰强烈的抢救优先的优化情景中, 90.7%的保护对象栖息地覆盖范围有明显提升。三个优化情景中优先区的分布均可确定樟江主河道两岸、爽姑河上游、水令河上游等重点保护空缺地区, 亟待新建自然保护地, 强化保护管理措施。樟江流域生物多样性保护空间格局优化研究对互补算法保护规划具有典型意义, 对我国乃至世界上这一独特的喀斯特资源及其所提供的生态系统服务具有重要保护作用和科学支撑。
[1] RANDS M R, ADAMS W M, BENNUN L, et al. Biodiversity Conservation: Challenges Beyond 2010[J]. Science, 2010, 329(5997): 1298–1303.
[2] BRUGIERE D, SCHOLTE P. Biodiversity gap analysis of the protected area system in poorly–documented Chad[J]. Journal for Nature Conservation, 2013, 21(5): 286–293.
[3] BUTCHART S H, WALPOLE M, COLLEN B, et al. Global Biodiversity: Indicators of Recent Declines[J]. Science, 2010, 328(5982): 1164–1168.
[4] KULLBERG P, MOILANEN A. How do recent spatial biodiversity analyses support the convention on biological diversity in the expansion of the global conservation area network[J]. Natureza & Conservacao, 2014, 12(1): 3–10.
[5] RODRIGUES A S, BROOKS T M. Shortcuts for Biodiversity Conservation Planning: The Effectiveness of Surrogates[J]. Annual Review of Ecology, Evolution, and Systematics, 2007, 38(1): 713–737.
[6] DE ALBUQUERQUE F S, BEIER P. Using abiotic variables to predict importance of sites for species representation[J]. Conservation Biology, 2015, 29(5): 1390–1400.
[7] Moilanen A, Wilson K A, Possingham H p. Spatial conservation prioritization: quantitative methods & computational tools[M]. Oxford University Press, New York, 2009.
[8] ROBERTS C M, MCCLEAN C J, VERON J, et al. Marine biodiversity hotspots and conservation priorities for tropical reefs[J]. Science, 2002, 295(5558): 1280–1284.
[9] RAMIREZ F, AFAN I, DAVIS L S, et al. Climate impacts on global hot spots of marine biodiversity[J]. Science Advances, 2017, 3(2).
[10] VEACH V, MININ E D, POUZOLS F M, et al. Species richness as criterion for global conservation area placement leads to large losses in coverage of biodiversity[J]. Diversity and Distributions, 2017, 23(7): 715–726.
[11] MOILANEN A, POUZOLS F, MELLER L, et al. Zonation: spatial conservation planning and software v. 4 [EB/OL]. http: //cbig. it. helsinki. fi, 2014.
[12] WATSON J E, DUDLEY N, SEGAN D B, et al. 2014. The performance and potential of protected areas. Nature [J], 515: 67–73.
[13] MA Z, CHEN Y, MELVILLE D S, et al. Changes in area and number of nature reserves in China[J]. Conservation Biology, 2019, 33(5): 1066–1075.
[14] ELITH J, FRANKLIN J 2013. Species distribution modeling [M], Encyclopedia of Biodiversity: Second Edition. Elsevier Inc. : 692–705.
[15] 马琳, 李俊清. 基于系统保护规划的长白山阔叶红松林保护网络优化研究[J]. 生态学报, 2019, 39(22): 8547– 8555.
[16] 梁晨, 李晓文, 崔保山, 等. 中国滨海湿地优先保护格局构建[J]. 湿地科学, 2015, 13(6): 660–666.
[17] LEHTOMäKI J, KUOKKANEN P, HANSKI I, et al. Planning of forest conservation areas using high–resolution GIS data and software for spatial conservation prioritization [J]. Forest Ecology and Management, 2009, 258: 2439–2449.
[18] MOILANEN A, ANDERSON B J, EIGENBROD F, et al. Balancing alternative land uses in conservation prioritization [J]. Ecological Applications, 2011, 21: 1419–1426.
[19] 史雪威, 张路, 张晶晶, 等. 西南地区生物多样性保护优先格局评估[J]. 生态学杂志, 2018, 37(12): 3721–3728.
[20] ALBUQUERQUE F, BEIER P. Rarity–Weighted Richness: A Simple and Reliable Alternative to Integer Programming and Heuristic Algorithms for Minimum Set and Maximum Coverage Problems in Conservation Planning[J]. PloS one, 2015, 10: 1–7.
[21] ALBUQUERQUE F, BEIER P. Global patterns and environmental correlates of high–priority conservation areas for vertebrates[J]. Journal of Biogeography, 2015, 42: 1397–1405.
[22] ASTUDILLO–SCALIA Y, ALBUQUERQUE F. Why should we reconsider using species richness in spatial conservation prioritization[J]. Biodiversity and Conservation, 2020: 1–13.
[23] 徐建英, 陈利顶, 吕一河, 等. 保护区与社区关系协调: 方法和实践经验[J]. 生态学杂志, 2005, 24(1): 102–107.
[24] 刘洋, 吕一河, 陈利顶,等. 自然保护区生态旅游影响评价: 进展与启示[J]. 自然资源学报, 2005,20(5): 771–779.
[25] GAME E T, KAREIVA P, POSSINGHAM H P, et al. Six common mistakes in conservation priority setting[J]. Conservation Biology, 2013, 27(3): 480–485.
Biodiversity-based spatial optimization for natural conservation areas of Zhangjiang River basin
FU Wei*, ZHEN Guangrun
School of Architecture and Urban Planning, Beijing University of Civil Engineering and Architecture, Beijing 100044, China
It assumes an essential role to optimize the spatial pattern of ecological conservation within the scope of ecological civilization construction in China, which is also the frontier orientation of international research regarding ecological conservation. Assessing resource endowments, identifying and prioritizing and predicting conservation areas are critical to the protection and management of endangered species. Although conservation areas account for 11.9% of the Zhangjiang River watershed, significant habitat degradation has occurred across 77.9% of the watershed, highlighting an urgent need to balance development and protection. Based on a complementary non-exhaustive algorithm, the Zonation4GU software can produce ideal outcomes in optimizing existing conservative networks and characterizing rarity and uniqueness. This paper presents a study of the route towards spatial integration and optimization in natural conservations in the Zhangjiang River watershed based on the software. First, the distributional patterns of habitats of species, weights of species protection and planning units were manually preprocessed; the layer of conditions for imported habitat degradation and the layer of protection cost were iteratively selected; finally, optimization solutions to three different scenarios were proposed in combination of the artificial interference. The results show that the area prioritizing protection has an ideal coverage on habitats of proxy species, optimization solutions to scenario prioritizing ecological integrity and that prioritizing the minimum social, economic and land-resource costs both present as tire-1 priority areas with an construction expansion rate smaller than 8% and all protected targets receiving protection; after around 27% of the all priority areas were incorporated, the two optimization solutions exhibited a 70.0% and 59.0% rise, respectively, in covering the habitats of all protected targets. Although the optimization scenario prioritizing ecological integrity produces better performances than that prioritizing the minimum social, economic and land-resource costs, it entails a higher cost of protection. With respect to the optimization scenario prioritizing intense artificial interference such as urgent conservation, 90.7% of the inhabits for the protected targets have seen a significant coverage increase. From the technical and operative perspective, this study provides a viable route towards creating an integration and optimization solution to China’s natural conservation system.
systematic conservation planning; conservation gap; Karst region; integrated watershed protection; conservation targets
10.14108/j.cnki.1008-8873.2022.01.006
傅微, 甄广润. 基于生物多样性的樟江流域自然保护地空间优化[J]. 生态科学, 2022, 41(1): 50–58.
FU Wei, ZHEN Guangrun, SUN Zhe, et al. Biodiversity-based spatial optimization for natural conservation areas of Zhangjiang River basin[J]. Ecological Science, 2022, 41(1): 50–58.
K903
A
1008-8873(2022)01-050-09
2020-05-21;
2020-07-10基金项目:国家自然科学基金项目(41901220) ;北京市优秀人才培养资助青年骨干个人项目(24269220006)
傅微(1988—), 女, 湖南长沙人, 博士, 讲师, 主要从事景观生态规划研究, E-mail: fuwei0807@163.com
傅微
猜你喜欢
旅游学刊(2022年5期)2022-05-31
先锋(2019年7期)2019-09-09
绿色中国(2018年11期)2018-11-06
汉语世界(The World of Chinese)(2018年3期)2018-10-22
厦门航空(2018年4期)2018-04-25
海峡姐妹(2017年12期)2018-01-31
语文世界(初中版)(2017年5期)2017-06-22
作文与考试·初中版(2017年12期)2017-04-19
西南学林(2011年0期)2011-11-12
作文大王·中高年级(2008年12期)2008-12-19
