
铁冬青叶绿体全基因组及系统进化分析
2022-02-18 12:21黄红宝何应会黄欣孙开道秦波黄耀恒
农业研究与应用 2022年5期
黄红宝,何应会,黄欣,孙开道,秦波,黄耀恒
(广西壮族自治区林业科学研究院/广西特色经济林培育与利用重点实验室,广西南宁 530002)
冬青属(Ilex)隶属冬青科(Aquifoliaceae),包含近700 个种[1],属下有3 个亚属,即冬青亚属(Subgen.Ilex),多核冬青亚属(Subgen.Byronia)和落叶冬青亚属(Subgen.Prinos)。其中冬青亚属包含的种较多,因而其又被划分为若干个组,而每组下面又被划分为若干个系[2]。中国分布有丰富的冬青亚属植物资源,有研究表明在中国范围内已被发现和鉴别的冬青亚属植物超过200 种[3,4]。由于近缘种在形态上的高度相似性,仅仅依靠形态特征去鉴别它们是困难且容易引起混乱的。近些年,基于叶绿体全基因组的系统发育学在大量植物中得以应用。相较于核基因组,叶绿体基因组多为母系遗传,具有结构保守稳定、进化速率适中等特点,能较好的提供物种进化信息[5]。同时,随着越来越多的植物叶绿体基因组数据被提交到NCBI(National Center for Biotechnology Information)数据库,数据的充实将进一步增强叶绿体基因组比较方法的科学性和真实性。
铁冬青(Ilex rotunda)又名救必应、熊胆木等,原产广西、广东、福建、浙江等地,为冬青科常绿乔木或灌木[2]。铁冬青具有多种应用价值:一是材用价值,它的木材结构紧密,具有香气,可用作制作器具;二是观赏价值,铁冬青成熟植株枝繁叶茂、树型美观,可用作行道树和庭荫树,在结果期还可营造“满树红果”的景观;三是药用价值,它的茎皮、根叶可入药,在临床上常应用于抗炎抑菌、抗血栓等。为了明确铁冬青的系统进化地位,本研究通过高通量测序获得了铁冬青叶绿体基因组的完整序列,分析其基本特征,并通过构建系统进化树最终确定铁冬青的系统位置,其研究结果可为冬青科种质资源的鉴定,叶绿体基因组的利用及后续研究等提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为铁冬青,植株位于广西壮族自治区桂林市全州县(26°09’N,110°51’E)。2021 年11月采集新鲜的嫩叶,用餐巾纸擦拭表面后,放入硅胶中干燥保存。剪取铁冬青的枝叶制作凭证标本(编号为TDQ20211107),并保存于广西林业科学研究院标本馆中。
1.2 试验方法
1.2.1 基因组DNA 提取及测序
将干燥后的样品使用改良的CTAB 法[6]提取铁冬青基因组总DNA,质检合格后,采用Illumina Novaseq 6000 测序平台测序其叶绿体基因组。
1.2.2 叶绿体基因组组装和注释
经测序得到Raw reads 数据后,利用NGS QC Tool-Kit 软件[7]过滤低质量序列,获得Clean reads数据;使用SPAdes 软件[8]进行从头组装得到叶绿体全基因组;使用PGA 工具[9]注释基因组;最后利用OGDRAW[10]软件绘制基因组物理图谱。
1.2.3 叶绿体基因组特征分析
利用MISA 软件[11]对铁冬青叶绿体基因组进行SSR 位点分析,核苷酸重复次数依次设置为:8、5、4、3、3、3,两个SSR 位点间最小长度为100 bp,其余保持默认值。
1.2.4 系统发育分析
从NCBI 数据库下载总计34 个物种的叶绿体基因组,其中31 个物种来自冬青科(Aquifoliaceae),2 个物种来自青荚叶科(Helwingiaceae),1 个物种来自毛柴木科(Pennantiaceae),物种拉丁名及叶绿体基因组登录号详见表1。利用BLASTn(https://blast.ncbi.nlm.nih.gov/Blast.cgi)从每个物种的叶绿体基因组中挑选68 个共有的同源蛋白基因,并使用MAFFT 软件[12]比对氨基酸序列;接着利用PhyloSuite 软件ModelFinder 选择TVM+F+I+G4 模型[13];最后使用IQtree 2.0 构建最大似然(ML)系统发育树[14],Bootstrap 的值为1000,并选择Pennantia cunninghamii(NC_049 885.1)作为外类群。

表1 34 个物种拉丁名及叶绿体基因组登录号
2 结果与分析
2.1 叶绿体基因组基本特征
通过高通量测序和叶绿体基因组物理图谱构建的结果显示,铁冬青叶绿体全基因组为共价闭合的双链环状分子(图1),全长157,778 bp,其中IRS(Inverted repeats)长度为26,124 bp,LSC(Large single copy)长度为87,092 bp,SSC(Small single copy)长度为18,438 bp,全基因组GC 含量为37.70%(表2);共编码131 个基因,其中蛋白编码基因86 个、rRNA 基因8 个、tRNA 基因37 个(表3)。获得的铁冬青叶绿体全基因组序列已上传至NCBI,序列号为OL396585。

表2 铁冬青叶绿体基因组基本特征

表3 铁冬青叶绿体全基因组序列的特征值
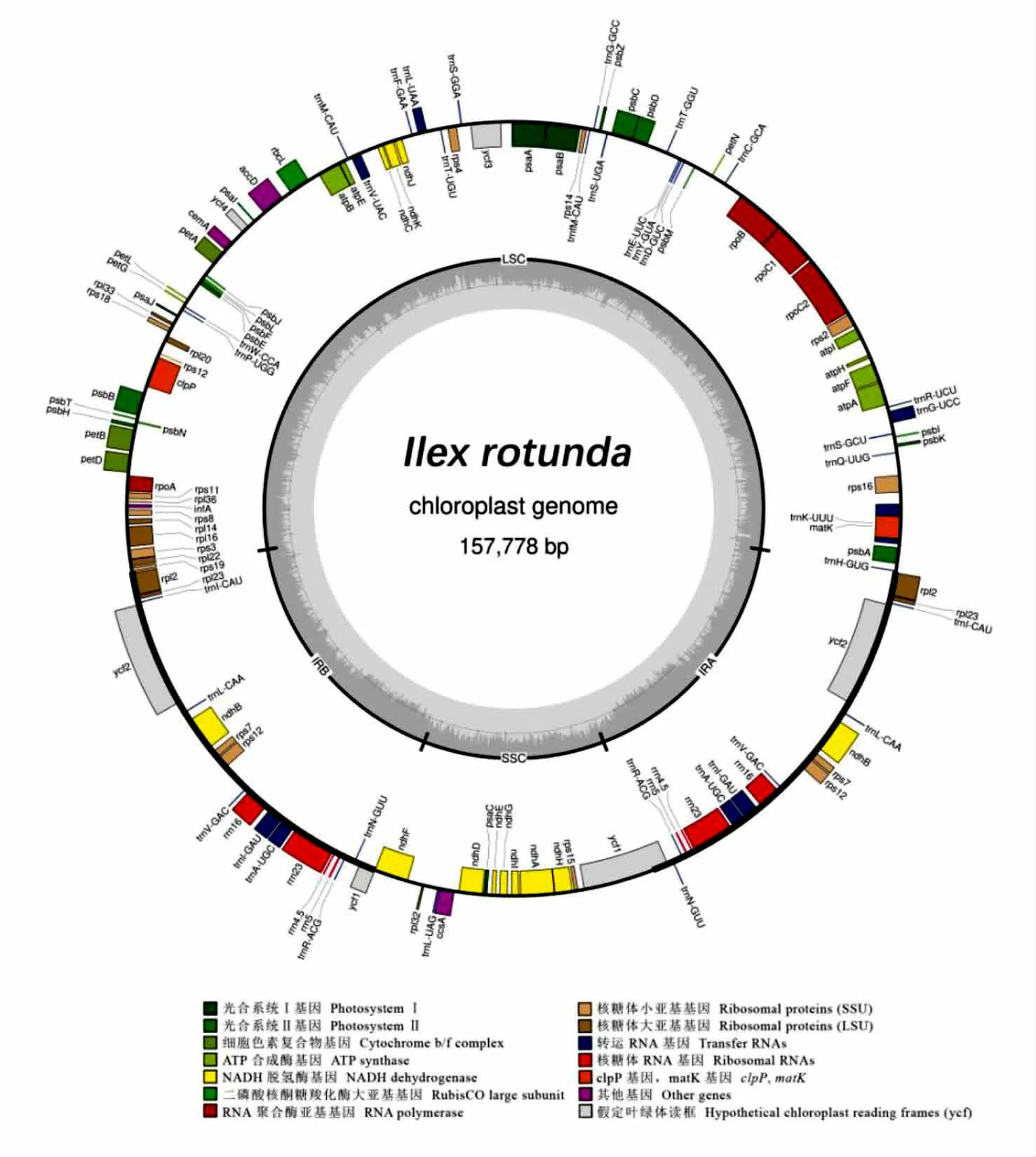
图1 铁冬青叶绿体基因组物理图谱
2.2 SSR 特征分析
在铁冬青叶绿体基因组中共检测到131 个SSR,其中大多数为单核苷SSR,共120 个(占91.60%),其中A/T 重复117 个,C/G 重复3 个;二核苷酸SSR 4 个(占3.05%),为AT/AT 重复;三核苷酸SSR 1 个(占0.76%),为AAT/ATT 重复;四核苷酸SSR 4 个(占3.05%),其中AAAG/CTTT 重复1 个,AAAT/ATTT 重复3 个;六核苷酸SSR 2 个(占1.53%),均为AATACT/AGTATT 重复(表4)。

表4 铁冬青叶绿体全基因组SSR 信息
2.3 系统进化分析
基于34 个物种叶绿体基因组共有的同源蛋白(68 个)信息,构建了最大似然(ML)系统发育树。聚类结果显示(图2),本研究的铁冬青测试样品(OL396585)与已在NCBI 公开发表的铁冬青(MT764244.1)聚在一起,支持率为100%;且铁冬青隶属于冬青亚属单序冬青组(Sect.Lioprinus),与毛冬青(I.pubescens,KX462467.1)和网脉多脉冬青(I.polyneura,KX462468.1)亲缘关系较近。

图2 基于叶绿体基因组同源蛋白的34 个物种系统发育树
3 讨论与结论
本研究的铁冬青样品叶绿体全基因组长度为157,778 bp,与在NCBI 上已发表的铁冬青其他样品叶绿体基因组NC_056200.1(157743 bp)、MZ561473.1(157776 bp)、MT764244.1(157 780 bp)长度接近,同一物种4 个不同样品的叶绿体全基因组长度差异范围为2~37 bp;不同样品基因组长度上的差异可能是由于不同地理种源的遗传差异引起的。
SSR 标记具有共显性和高多态性[15],已作为一种可靠的分子标记被应用于植物群体遗传学[16]、种质鉴别[17]、遗传图谱构建[18,19]等研究中。本研究分析鉴定出铁冬青叶绿体全基因组种共含有131 个SSR,其中单核苷SSR 的数量为120 个,占总数的91.60%。丰富的SSR 位点可用于后续开发高多态性的SSR 标记引物,并作为铁冬青种内遗传多样性研究和物种鉴别的工具。
通过构建系统发育树能将分子数据应用于研究物种间发育关系。系统发育分析显示,本研究的铁冬青测试样品与NCBI 上已发表的铁冬青MT764244.1 聚在一起(支持率为100%),表明物种鉴别及其鉴别方法正确;其中铁冬青与近缘种毛冬青和网脉多脉冬青(I.polyneura)聚在一起,表明它们具有较近的亲缘关系和类似的物种起源,这与Chen 等[2]利用叶绿体基因组全长序列研究所得出的结论一致。研究结果进一步确定了铁冬青的系统位置,同时也印证了基于叶绿体全基因组的系统发育分析方法的可行性和可靠性。此外,系统发育树分析结果进一步显示,30 个冬青科植物分为分支Ⅰ和分支Ⅱ两大支(Clade),分支Ⅰ含有14 个物种,包括9 个传统分类方法的冬青亚属(Subgen.Ilex)物种及5 个“未确定(Uncertain)”物种;分支Ⅱ含有16 个物种,包括11 个传统分类方法的冬青亚属物种和5 个落叶冬青亚属(Subgen.Prinos)物种;即同属于冬青亚属的物种在系统发育树中分属于不同的两大分支(Clade)。可见,基于冬青科叶绿体基因组的系统发育结果与传统的分类研究结果出现了不一致的情况;而在Alexandra 等利用分子标记开展的冬青科系统发育研究中也出现了类似的结果[20],其具体的原因有待于深入研究。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
中华建设(2020年5期)2020-07-24
少儿科技(2019年9期)2019-09-10
小学生必读(低年级版)(2018年6期)2018-09-08
中成药(2018年7期)2018-08-04
——致坚守奋斗的你
音乐天地(音乐创作版)(2017年1期)2017-02-12
中央民族大学学报(自然科学版)(2015年3期)2015-06-11
