
作物从头驯化策略的提出与进展
2022-02-16 06:29:36张静昆李文佳曾鹏孟祥兵余泓李家洋
中国农业科技导报 2022年12期
张静昆 , 李文佳 , 曾鹏 , 孟祥兵 , 余泓 , 李家洋 ,3*
(1.中国科学院遗传与发育生物学研究所,北京 100101; 2.中国科学院大学,北京 100049; 3.海南省崖州湾种子实验室,海南 三亚 572025; 4.海南大学三亚南繁研究院,海南 三亚 572024)
种子是农业的“芯片”,育种技术的发展对于保障世界粮食安全发挥了重要的作用。从近代农业的发展历程来看,育种技术的创新使得育种效率不断提高,新型品种的更新换代使得粮食产量大幅提升,全球粮食总产量从1960年的8.63亿t增长到了2020年的29.96亿t。然而,到2050年世界人口将超过90亿,粮食产量需要再增加约60%才可以满足需求[1]。同时,气候变化等因素严重影响粮食产量,极端气候发生频率逐年升高。这些新的问题对于现有种质与传统育种技术提出了新的挑战。未来,培育高产优质且环境适应能力强的“智能作物”(smart crops)被认为是应对挑战的有效策略[2]。从头驯化(de novodomestication)策略可利用更为广泛的自然资源来培育全新类型的智能作物,有望引领新一轮的农业革命。本文综述了从头驯化的发展和技术路线、野生资源基础、驯化机制解析及底层生物技术,分析了从头驯化策略可能面临的挑战,旨在为其在作物育种上的广泛应用提供参考。
1 从头驯化策略简介
1.1 从头驯化的基本概念
从头驯化是指筛选具有特定优异性状(如耐盐碱、抗病虫、环境适应能力强、生物量大、能够合成重要天然化合物等具有重要应用前景的性状)的野生或半野生植物,综合应用基因组编辑、功能基因组学、生物信息学、染色体工程、合成生物学等技术与理论,高效精准改良野生植物的未驯化性状,使其能够适用于现代农业生产,从而培育出全新作物的一种育种策略[3]。所培育出的新型作物既保留野生材料的优势,又符合现代农业生产要求,大大丰富农业生态多样性。按照驯化物种的不同,特别是野生物种的驯化程度及与栽培作物是否存在生殖隔离等特征的差异,从头驯化路径大体可以分为3个主要方向(图1):①现有栽培品种的祖先野生植物的重新驯化,可快速地将自然驯化中丢失的抗逆性状导入到栽培品种中,拓宽现有栽培品种的遗传多样性,填补栽培品种缺乏的优异性状;②新野生植物的从头驯化,可对未被人类驯化过的优异野生物种进行快速从头驯化,突破现有作物的物种限制,培育全新作物类型;③加速半野生植物或孤生作物的驯化,可针对只适宜特定区域种植而育种投入相对较小的特色作物,快速改良其未驯化性状,助力发展地区特色农业。
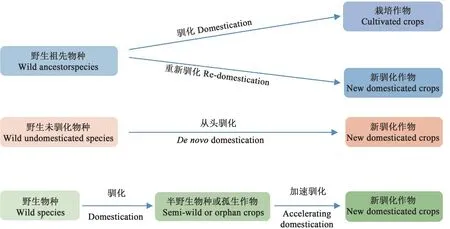
图1 从头驯化的主要策略Fig. 1 Major strategies of de novo domestication
1.2 从头驯化理念的提出
随着功能基因组学的快速发展,驯化事件的遗传机制被研究的越来越清晰。2013年,中国科学院植物研究所桑涛研究员和中国科学院遗传与发育生物学研究所李家洋院士综述了谷物作物驯化事件的分子遗传学基础,并提出通过科学的杂交设计可以加速作物驯化综合特征的遗传改良(speed up domestication syndrome),且随着遗传学、基因组学研究和先进的生物学技术的发展,此过程将进一步加速[4]。同年,中国科学院植物研究所桑涛研究员和葛颂研究员总结了栽培稻的驯化历程,提出数量性状位点定位、全基因组关联分析和基因组扫描等技术是挖掘栽培和野生物种有价值等位基因的有效手段,且利用广泛搜集的种质以及产生的等位基因库,结合分子育种可能会触发水稻品种的重新驯化(re-domestication)[5]。与此同时,以CRISPR/Cas9为基础的高效基因组编辑技术在植物和作物上首次应用,大大拓展了对植物基因组进行改造的技术手段[6-8]。2017年,巴西圣保罗大学 Lázaro Eustáquio Pereira Peres团队提出,可以利用最新的基因组编辑技术从头驯化具有抗逆性或其他目标性状的野生近缘种,并具体举例可以通过编辑野生多腺番茄(Solanum galapagense,可耐受高达70%的海水盐度)中的SP、FW2.2、FAS、LN、Cyc-B和S基因,有望创制出株型与果实性状均改良的高抗盐番茄[9]。2017年,丹麦哥本哈根大学的Michael Palmgren团队提出加速驯化(accelerating domestication)策略,并建议从模拟驯化事件和利用基因组编辑技术两大方面来实现并加速驯化[10]。
1.3 从头驯化取得突破性进展
2018年,Nature Biotechnology杂志同期发表了2篇文章,中国科学院遗传与发育生物学研究所高彩霞团队和许操团队合作,利用多基因组编辑技术精准靶向目标基因的编码区,顺式作用元件以及上游开放阅读框区域,创制出综合性状改良的番茄,同时又保持了野生番茄的天然抗性[11];Lázaro Eustáquio Pereira Peres团队和德国明斯特大学Jörg Kudla团队合作,通过对番茄祖先种野生醋栗番茄(Solanum pimpinellifolium)中控制重要农艺性状的主效基因进行编辑,快速实现了产量和品质优良性状的聚合,这两项重要的工作首次实现了野生种质的快速驯化,证明了从头驯化育种策略的可行性[12]。同年,美国冷泉港实验室Zachary B. Lippman 团队对孤生作物(orphan crop)灯笼果(Physalis pruinosa)进行驯化,改良了灯笼果的株型、挂果数量和果实大小[13]。
2021年,李家洋团队提出异源四倍体野生稻的从头驯化育种策略,该策略以南美起源的未被人类驯化过的四倍体野生稻为基础,利用其所具有的生物量大、耐逆性好、环境适应能力强的优势,提出了一种培育高产、抗逆、环境适应性强的全新粮食作物的方案,并突破了包括遗传转化在内的一系列四倍体野生稻快速驯化的技术瓶颈,成功对多个驯化性状进行快速改良,证明了该策略高度可行,有望突破现有栽培稻的育种瓶颈,为创制全新粮食作物、保障世界粮食安全开辟了新的路径[14]。
2 野生植物从头驯化的理论与技术基础
2.1 从头驯化技术路线
目前,从头驯化的整体技术路线已经得到比较完整的阐述[14],可分为4个阶段:①收集并筛选综合性状最佳的目标种质资源;②建立目标种质从头驯化技术体系,其中包括3个核心点,即高效遗传转化体系、高效基因组编辑技术体系和高质量参考基因组的建立;③品种分子设计与快速驯化,挖掘野生种质资源基因并确定功能基因,利用多基因编辑技术进行优良性状的聚合,评估综合性状;④新型作物的审定、推广与应用(图2)。
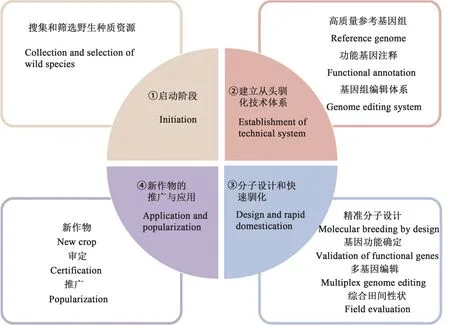
图2 从头驯化策略的实施路线Fig. 2 Practical route of de novo domestication
2.2 野生植物的资源基础
现代栽培种的遗传背景狭窄,已成为种质创新的重要瓶颈。野生种质资源通常具有抗病虫、耐盐碱等天然优势,且由于未被大面积单一化种植,遗传多样性丰富。因而,利用野生种质鉴定克隆新基因或基因模块已成为育种改良的重要方向之一。例如将野生龙葵(Solanum americanum)的Rpi-amr1基因导入马铃薯中,对19种可导致马铃薯晚疫病的致病疫霉具有抗性作用,为防治马铃薯晚疫病提供全新遗传资源[15]。同时,基于野生种质构建的遗传群体和等位基因库可以用来解析优异等位驯化的起源与演变规律,为作物从头驯化提供候选基因。虽然从野生资源中已经鉴定出了一些控制抗病虫、耐盐碱、抗除草剂、营养高效等重要性状的基因,但整体由于野生资源难与栽培品种杂交或染色体重组效率很低,能够真正应用到农业生产的仍然非常有限。
种质资源作为基础材料,从搜集、整理、鉴定、扩繁、记录、存储和利用都要进行有效保护与科学研究。广泛搜集不同起源生境的种质,建立高通量的表型鉴定检测平台,系统地对这些资源进行鉴定,筛选具有优异性状的种质资源,是野生资源利用以及从头驯化的重要基础。我国野生资源非常丰富,目前收集并保存的农作物种质资源合计超过52万份,但仅有10%的资源材料有初步的表型鉴定记录[16]。如何建立特定育种目标的筛选体系,如抗病、抗虫苗圃以及不同浓度的盐池等,进行规模化筛选和优异野生资源鉴定,将对从头驯化研究具有重要意义。同时,种质的利用要遵守相应的法律法规,保障种质的合理输出与输入。
2.3 从头驯化的功能基因组学基础
作物驯化是人类文明起始的里程碑事件。作物驯化的本质是人类对具有潜力的野生植物进行选择,从大量的自然变异中保留优异的等位基因,使得植物性状发生改变以适合农业生产,满足人类需求。在漫长的作物驯化中,作物落粒性丧失、种子变多、休眠性下降、果实增大、株型直立。此外,种子颜色、穗型、生育期等都发生改变[17]。栽培品种与野生祖先种中表现出的性状差异被称为“驯化综合特征”。Meyer等[18]研究表明,在203种作物中,每种作物平均包含有2.8个驯化综合特征,84%的作物具有2~5个驯化综合特征,而有些作物则多达7个。然而,随着作物驯化与育种,优异品种被大面积单一化种植,不可避免地降低作物遗传多样性。从头驯化策略是从野生或半野生物种的天然优异特性和遗传多样性出发,针对驯化综合特征进行设计和定向改良。随着遗传学和功能基因组学的发展,控制驯化性状形成的特定基因相继得到克隆[19]。而作物驯化和改良相关理论知识也是对野生植物进行从头驯化的关键基础与前提。下面总结了重要作物关键驯化性状的遗传基础。
亚洲栽培稻(Oryza sativa)是由亚洲起源的二倍体普通野生稻(Oryza rufipogon)驯化而来。目前,从野生稻到栽培水稻的主要驯化性状及其基因已经得到了较为清楚的解析。野生稻落粒性的丧失是由于qSH1基因和Sh4基因分别在调控区和编码区发生的突变导致[20-21]。水稻由匍匐生长变为直立生长是由PROG1基因的突变导致[22-23]。穗型由松散变为紧凑的分子基础是OsLG1上游3.3 kb 调控区 SNP 的突 变或其上 游 10 kb 的SPR3位点变异[24-25]。An-1、An-2/LABA1、RAE2分别编码螺旋-环-螺旋蛋白、细胞分裂素激活酶和表皮模式因子类蛋白EPFL1,均能正调控芒长,亚洲栽培稻中这些基因的功能丧失导致芒变短或无芒表型[26-28]。GW5/GSE5启动子区域1.2 kb的缺失会抑制该基因的表达,从而促进颖壳细胞的分裂,增加栽培稻的粒宽与粒重[29]。Rc是控制种子颜色的主效基因,其第6个外显子上14 bp的缺失不仅可以调控种子中原花色素的合成,也促进ABA合成基因的表达,同时减弱栽培稻的种子休眠性[30-31]。Bh4第3外显子上存在22 bp的缺失致使栽培水稻谷壳颜色变为黄色[32]。同时,品质性状也在驯化选择中得到提升。Wx基因编码颗粒淀粉合成酶,能够影响蒸煮品质、胶稠度以及糊化温度,已报道的Wx基因至少存在8种复等位变异,且具有地域性[33]。除以上驯化基因外也有许多控制重要农艺性状的基因在现代作物育种中发挥重要作用,如绿色革命基因SD1、理想株型基因IPA1、分蘖角度基因TAC1、穗粒数基因Gn1a和FZP、落粒基因SH5和SHAT1、休眠基因Sdr4和OsG,以及粒长基因GS3、GL3.1、GL7等。此外,作物生育期决定了种质扩散与适宜的地域分布,DTH7、Ghd7、Ghd8、HD1等基因的不同等位组合共同决定了水稻品种的感光性和地域适应性[34-35]。
玉米(Zea maysL.)是从墨西哥和中美洲起源的祖先种大刍草(Purus frumentum)驯化而来,研究人员利用玉米和大刍草杂交群体,明确了玉米驯化中5个重要的性状发生了改变,分别是雌穗花序结构、穗行数、穗轴脱落性、籽粒裸露性以及分蘖数目[36-38]。穗花序形态建成基因RA1控制腋生分生组织的命运,其开放阅读框位置发生氨基酸种类和数目的突变而降低了RA1的表达水平,进而降低雌穗分支[39]。ZFL1和ZFL2外显子区域的功能缺失突变使玉米果穗花序为多列式,通过增加穗行数而提高产量[40]。ZmSh1-1、ZmSh1-5.1和ZmSh1-5.2是控制果穗脱落的基因,3个基因均发生结构变异导致功能丧失,且Sh1在高粱、水稻中受到平行选择[41]。TGA1基因编码SBP转录因子,其氨基酸突变能使籽粒从坚硬稃壳包裹中裸露出来[42]。TB1编码TCP转录因子,玉米中该基因上游60 kb区域的转座子插入能够增强TB1的表达,进而抑制分蘖数目,同时增强顶端优势[43]。
小麦(Triticum aestivum)是在中东新月沃地驯化而来。野生小麦具有碎穗性状和包壳性状,从而便于种子扩散和趋避鸟类。小麦TtBtr1-A外显子中2 bp的缺失和TtBtr1-B外显子中4 kb的插入分别导致TtBtr1-A和TtBtr1-B蛋白功能丧失,使小麦种子不再破碎,从而提高收获指数[44]。在栽培小麦中Q基因的SNP突变使得Q表达上调,从而籽粒变得容易脱落。Q基因受到明显选择且控制着小麦多个驯化性状,如颖片形状和坚韧性、穗轴韧性、穗长、株高和抽穗时间等[45-46]。Tg1-B和Tg1-D也控制小麦脱粒性状[47]。春化特性直接影响小麦的种植范围,其主要受春化基因Vrn1调控,该基因编码MADS转录因子,能促进开花;Vrn1启动子或内含子的变异决定了其表达水平,春小麦中Vrn1基因表达量较高,而冬小麦中Vrn1表达量处于较低水平[48-49]。
番茄驯化同样改良了株型和果实大小。野生番茄为无限生长型,营养生长和生殖生长交替进行。SP基因编码抗花素,该基因1个氨基酸的突变使栽培番茄株型矮化紧凑,坐果集中且适于密植[50]。抗花素基因在大豆、草莓、向日葵等驯化改良中同样得到应用[51]。fw2.2基因顺式区域的改变降低其表达,进而增加了果实大小[52]。此外,腔室数目的增加被认为是番茄果实驯化中最重要的增产因素,fasciated基因ATG下游330 bp处7 bp的缺失和第1个外显子6~8 kb的插入使得fasciated表达下调从而大大增加果实中腔室数目[53]。随着研究的深入,作物驯化不仅仅使植物在可见的株型等性状上发生了改良,在代谢层面也造成了很大变化[54-55]。在果实作物中,风味物质、色素等次生代谢物的变化是高度常见的驯化事件,且发生频率远高于其他农艺性状改变[18],可能是由于它们对果实品质有决定性影响。
近年来科学家发现,尽管驯化的物种、地点不同,人类对不同作物的许多性状进行了趋同驯化(convergent domestication)。例如玉米ZmKRN2基因5’UTR的变异在玉米中受到驯化选择,降低了该基因的表达量,导致穗行数增加,而水稻中的同源基因OsKRN2也位于驯化选择区域内,功能丧失可以增加水稻穗粒数[56]。玉米ZmTB1、水稻OsTB1、大麦HvTB1、小麦TaTB1均作为分蘗的负调控因子发挥功能[57]。除了性状上的趋同选择,在分子层面也有越来越多的研究表明,重要的基因在不同物种中存在平行驯化(parallel domestication),例如调控种子休眠的G基因在不同科植物(大豆、水稻、拟南芥等)驯化中受到平行选择[58]。控制生育期的主效基因HD1在高粱、谷子以及水稻中均受到平行选择,即使在不同谷物作物中受到选择的变异类型不同,但都改变了作物生育期和地域适应性[59]。此外,质体基因组是如何受到驯化选择仍需进一步研究。随着新技术的发展,更多驯化重要基因的克隆将为从头驯化策略提供可用的遗传资源。
2.4 从头驯化的基础技术
高效的遗传转化体系是对目标种质资源进行遗传操作的基础,也是从头驯化的基本底层技术。植物遗传转化技术经过不断发展,已产生了多种操作方法,其中农杆菌介导的遗传转化方法因具有高效、经济便捷、操作简单等优点,已在水稻、玉米、油菜、小麦、大麦、大豆、番茄、棉花、高粱、土豆、苹果、无花果等植物中被广泛应用,农杆菌介导的遗传转化方法也是目前进行植物基因组编辑的主要方法[60]。然而,尽管植物转基因技术已经得到不断优化与发展,但目前仅有约0.1%的植物能够进行遗传操作,如何在野生植物中进行遗传转化仍然是非常有挑战性的一项前沿工作[61]。以水稻为例,虽然世界上首例转基因水稻问世距今已有30多年时间,现在的遗传转化体系仍只能主要适用于亚洲栽培稻,特别是粳稻品种的转化效率较高,而籼稻品种仍只有特定遗传背景的品种可以转化[62-64]。对于野生稻而言,只有AA、BB、FF基因组的二倍体野生稻实现了低效率的遗传转化[65]。近期,CCDD异源四倍体野生稻以成熟胚作为外植体的组培再生和遗传转化取得重大突破,而这一技术瓶颈的突破也是开展异源四倍体野生稻从头驯化的前提[14]。未来在不同野生植物的从头驯化实践中,建立目标种质资源的遗传转化体系尤为重要。近几年研究发现一些发育调节因子可以提高遗传转化效率。如35S强启动子驱动ZmBbm1结合Nos弱启动子驱动Wus2的方法可以在单双子叶作物中诱导芽的从头再生,进而突破遗传转化的组织培养和基因型限制[66]。过表达GRF4及其共调节子基因GIF1可以促进小麦、黑小麦、水稻、柑橘愈伤组织的再生能力[67]。近期,朱健康院士团队开发出一种切-浸-萌芽(cut-dipbudding)的方法,无需进行组织培养,成功实现了多种植物(橡胶草、小冠花、甘薯、臭椿、辽东楤木、臭茉莉)的遗传转化和基因编辑[61]。此外,基因枪法、花粉管通道法及新型纳米材料等也能用于创制遗传改良材料。
基因组编辑可以按照设计快速创制新的等位基因,是从头驯化的关键技术。现有的基因组编辑技术包括单碱基定点编辑、单基因和多基因编辑、先导编辑等系统,在作物育种方面展示出强大的应用前景。尤其是CRISPR/Cas系统能够使用多种Cas核酸酶变体识别不同PAM,可以满足基因组靶点设计的需求。针对野生植物,进一步提升基因组编辑效率、扩大靶点设计范围、增加多靶点编辑效率、增加纯合体并降低嵌合体比例,都能极大地提高从头驯化的效率。在递送系统改良上,还可以利用RNA病毒,如烟草花叶病毒载体(TMV)和烟草脆裂病毒载体(TRV)系统地大量感染植物,并输送到植物的生长点,且病毒RNA基因组不会整合到植物基因组中,扩展了CRISPR/Cas9系统的实用性[68]。双生病毒载体整合CRISPR/Cas9系统在水稻中可实现外源基因的定点敲入[69]。对特定细胞器进行基因组编辑近年来取得了很大进展,例如基于TALEN系统对叶绿体、线粒体基因的编辑[70-71],是对质体基因组里潜在的驯化基因进行改造的有利工具。
基因组测序与组装是解码目标种质遗传信息的前提,驯化相关基因的序列信息为从头驯化中应该对哪些基因进行什么样的基因编辑提供了基础数据。自2000年首次报道拟南芥的完整基因组以来,截至2020年,已有788个植物物种被测序,其中有47个物种的基因组已组装到染色体水平[72]。随着测序成本的降低和样本通量的扩大,二代测序、实时荧光单分子测序、染色体构象捕获等技术的综合应用开启了植物基因组测序新篇章。一方面对于复杂基因组的组装也取得重要进展,在被子植物中,48种四倍体、5种六倍体和6种八倍体植物已完成测序[72]。这些参考基因组的绘制可以为目标种质的遗传改良提供基因资源。另一方面,许多作物的基因组学研究已经进入泛基因组时代。泛基因组的构建可以得到物种全面且准确的变异信息,例如对251份来自不同国家和地区的亚洲栽培稻、亚洲野生稻、非洲栽培稻和非洲野生稻构建高质量的水稻泛基因组,鉴定了抗逆基因家族NLRs图谱,解析了亚洲稻和非洲稻耐水淹适应性,揭示了落粒性丧失以及株型从匍匐到直立的平行驯化[73]。此外,已构建番茄、大豆、向日葵、油菜等泛基因组,有助于种质资源的挖掘和利用[74-76]。
2.5 从头驯化的相关政策
从头驯化产生的新作物如何能够真正地应用于农业生产,需要国家政策保障,这主要包含3个层面的保障。
第一层面,从头驯化以基因组编辑技术为基础,首先是健全完善对基因组编辑植物的管理政策。为规范农用基因组编辑植物生物安全评价工作,2022年1月24日,农业农村部印发的《农业用基因编辑植物安全评价指南(试行)》指出,对于未引入外源基因的基因组编辑植物,可依据分子特征、遗传稳定性、环境安全和食用安全等标准进行生物安全综合评价,申请生产应用安全证书。除了国内对基因组编辑育种的重视,国际上对基因组编辑技术的应用也进行扶持。美国是世界上基因编辑作物产业化领先的国家。2014年美国批准了首个基因组编辑油菜品种SU Canola(耐磺酰脲类除草剂)的商业化种植,之后又批准了150多种基因组编辑新植物[77]。2021年,美国发布的《面向2030年食品和农业的技术突破》(Science Breakthroughs to Advanced Food and Agricultural Research by 2030)中指出,应当鼓励突破性的基因组学和精准育种技术。法国已将基因组编辑作物视为非转基因植物,英国也表示将对基因组编辑作物采取更为宽松的监管方案。俄罗斯通过了《2019—2027年联邦基因技术发展规划》,指出主要目标为加速发展包括基因组编辑在内的基因技术。日本已对基因编辑农作物豁免转基因产品监管程序。
第二层面,国家要建立和完善对从头驯化作物的管理与审定政策。仅仅有政策支持,没有审定渠道,新种子就没办法到农民手里在田间生产。从头驯化新作物可能是之前种子法未涵盖的品种,对于新型作物的管理需要制定详尽的审定程序、相关条例,发布生产性实验和申请安全证书的相关规定与流程。确保生物安全前提下探索更为高效的审定模式,让相关产品有法可依进行推广。攻克生物技术育种产业化的障碍,激发科研机构、种子企业的创新动力,推动种业发展。
第三层面,考虑社会公众对于基因组编辑植物的接受程度。加大前期科普工作,避免公众对于基因组编辑作物缺乏科学的认识而阻碍新作物进入市场。政府在加强基因组编辑作物监管的同时,让生物安全与风险评估更加透明化,广泛听取公众意见。将科技创新与科普工作相结合,传播科学思想,进而真正促进编辑作物的商业化。
3 挑战与展望
种子是农业的芯片,种质创新在驱动农业发展中发挥着重要作用。从头驯化策略大大拓宽了人类可利用的遗传资源,开启了全新的作物育种方向。但要真正实现从头驯化作物,仍需多方面的努力:一是保护好种质资源的存储与扩繁,系统加强种子的收集、整理、存档、存储与扩繁等各个环节;二是解析作物驯化规律,揭示重要农艺性状形成的关键基因以及调控网络;三是开发高效多元的基因组编辑技术和遗传载体的递送系统,在通用的技术体系下,开发特定野生植物的遗传转化和递送系统;四是大量创制从头驯化新材料,生成全新的遗传资源库,筛选新型种质资源;五是探索并建立从头驯化新作物的相关政策法规,特别是从头驯化以基因组编辑为基础,需要健全基因组编辑品种的审定与管理办法,且涉及的物种可能是之前未涵盖的物种,需要完善种子法相关规定等;六是开发适宜的商业模式,做好作物推广以及科普工作,才能真正保障从头驯化新作物的商业化生产。这些工作上下游串联了多个学科,需要系统开展广泛深入的合作研究。
从头驯化育种策略有望引领育种新方向。野生或半野生植物中蕴含着栽培品种中已经丧失的优异基因资源。通过对野生植物的快速驯化可以培育出高抗作物,提高作物环境和气候适应性,减少农药等使用,发展环境友好型农业;选取能够在边际土地上生长的野生植物,有望实现边际土地的利用,增加可用的耕地面积;利用多倍体野生植物在生物量和环境适应力上的天然优势,结合杂种优势利用,有望能够突破现有作物产量瓶颈,大幅提高作物产量。在筛选目标种质时,选择具有以上优势的野生或半野生植物,可以培育出综合性状更加优良的新作物,为世界粮食安全保驾护航。
猜你喜欢
区域治理(2022年40期)2022-11-27 04:01:54
今日农业(2022年13期)2022-09-15 01:18:00
今日农业(2021年11期)2021-08-13 08:53:24
动漫界·幼教365(小班)(2019年10期)2019-10-28 02:04:20
动漫界·幼教365(大班)(2019年10期)2019-10-28 01:54:09
动漫界·幼教365(中班)(2019年10期)2019-10-28 01:53:17
中国麻业科学(2018年6期)2018-04-09 11:22:12
西南农业学报(2016年5期)2016-05-17 05:42:21
广西林业科学(2016年3期)2016-03-16 05:43:21
遗传(2014年3期)2014-02-28 20:58:49
