
花溪饮用水源保护区植物多样性特征及区系分析
2022-02-15 07:14:00相孟达聂雪婷汤晓辛
中国野生植物资源 2022年1期
杨 丹,相孟达,邓 伟,聂雪婷,张 潮,金 毅,汤晓辛
(1.贵州师范大学西南喀斯特山地生物多样性保护国家林业和草原局重点实验室,贵州贵阳550001;2.贵州师范大学贵州省植物生理与发育调控实验室,贵州贵阳550001;3.贵州师范大学生命科学学院,贵州贵阳550001)
物种多样性是指物种水平上的生物多样性,从生态学角度对物种的多样性及群落组织进行的研究[1],物种多样性不仅可以反映群落中物种的丰富度和均匀度[2],也能反映群落与环境因子的关系[3]。植物的物种多样性为人类生存提供丰富的物质资源和环境条件,是生态系统健康程度的重要指标,能够揭示生态系统变化、维持群落生产力[4],同时也是构成生态系统服务功能的重要部分之一[5]。研究群落中的物种多样性可以更好的认识群落的组成和发展状况[6],同时对环境的保护、恢复和预防珍稀濒危物种的丧失具有重要意义。
贵阳市花溪十里河滩国家城市湿地公园主要位于贵阳市花溪区中心城区的北部,在环境上属于亚热带湿润气候下的高原岩溶丘陵区,属长江水系,兼顾湿地公园的保护、观赏、文化和科普价值需要,区内水热条件优越,植物多样性比较丰富,为许多物种提供了理想的栖息场所。花溪水库为贵阳市重要饮用水源保护区,也是花溪湿地公园重要的组成部分,该水源地周边的生物多样性对于保障花溪区城乡居民用水安全和满足人民生产生活需要具有重要意义。对花溪湿地公园的调查目前有鸟类多样性及浮游植物的研究[7-8],但缺少该水源区域植被和物种多样性系统的调查研究。因此,以花溪水库至阿哈湖水库区域为研究对象,进行陆生植物的监测,对环境植被进行其区系组成与物种多样性的分析,对保护花溪水库饮用水源保护区具有重要意义,也为花溪湿地生态系统内珍稀动植物栖息地的保护及湿地生态系统的保护和恢复提供重要的基础资料。
1 研究地区与研究方法
1.1 研究区概况
贵阳市花溪区位于贵州高原中部、贵阳市的南部,地处东经106°27′~106°52′,北纬26°11′~26°34′。花溪水库位于南明河上游花溪河段,为山区雨源性河流,径流由降水补给,径流特性与降水特性基本一致,径流年内分配不均匀,洪、枯变化较大。下游距花溪仅3 km,距贵阳市市区仅20 km,坝址以上控制面积为315 km2,距贵阳市市区仅20 km。海拔高程多在1100~1300 m之间,多年平均气温15.0℃,年平均最低气温11.6℃,最高气温为19.4℃,多年平均风速2.2 m/s,多年平均相对湿度为82%,多年平均5~10月径流量占全年的80.2%左右,多年平均11月~次年4月月径流量占全年的19.8%左右,年最小径流量多出现在每年12月~次年2月份,花溪水库多年平均流量为5.14 m3/s,常年受西风带控制,属于亚热带湿润温和型气候。
1.2 研究方法
1.2.1 样地建设与调查方法
根据所调查区域覆盖范围的大小,设计样点进行植物调查。调查范围为花溪水库至阿哈湖水库区域,选取植被较为丰富的斜坡、山丘等区域,覆盖人为影响区域,以样地Ⅰ为调查起始位点,依次进行样地Ⅰ、样地Ⅱ、样地Ⅲ、样地Ⅳ、样地Ⅴ、样地Ⅵ、样地Ⅶ、样地Ⅷ,其详细的地理信息如表1。植被调查主要包括乔木、灌木、草本的植物种类、数量和生境,采取野外记录、拍照、采集标本和室内鉴定等方法。根据群落类型随机选取样方,根据各调查点生境特征,采取不同调查方法。8个样点的样方面积为乔木群落20 m×20 m、灌木林5 m×5 m以及草本1 m×1 m,记录物种数和株数。植物标本鉴定主要依据《中国植物志》、《中国高等植物图鉴》、《贵州植物志》、《中国蕨类植物科属志》等。区系成分分析主要参考《中国种子植物属的分布区类型》以及《世界种子植物科的分布区类型系统》。
1.2.2 植物区系分析
根据《中国植物志》在线数据库(http://frps.eflo‑ra.cn/),核对样地内出现物种的种名及其分类地位(表1);同时检索《中国湿地植被》(1999)分类原则,确定植被类型;区系成分分析以《中国种子植物区系统计分析》[9]和《种子植物分布区类型及其起源和分化》[10]、《中国蕨类植物科属志》[11]、《世界种子植物科的分布区类型系统》[12]、《中国种子植物属的分布区类型》[13]为依据,同时参照臧德奎对中国蕨类植物地理成分的划分方法[14],对样地内植物进行区系研究统计分析,在进行科属种的区系分析时,种下分类群均视作种级看待。
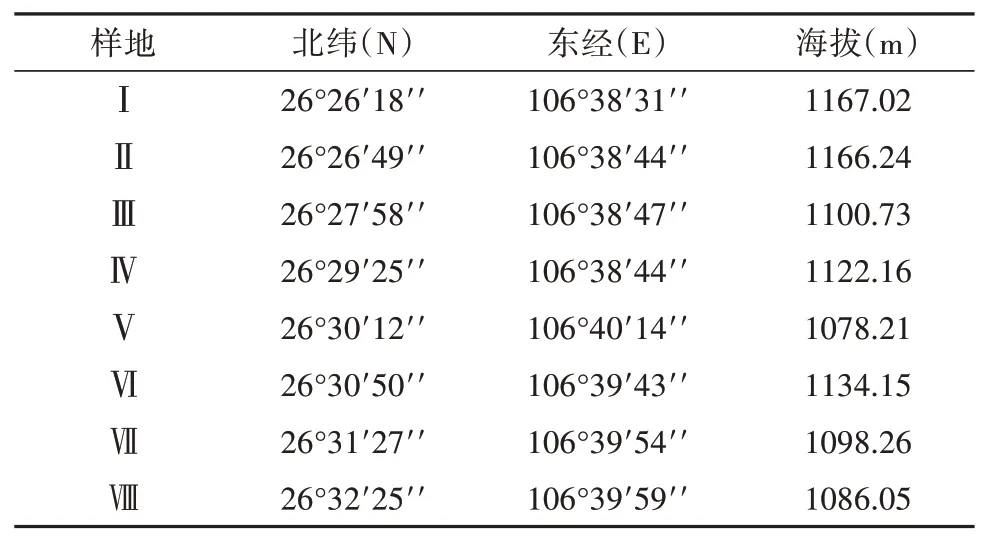
表1 陆生生物多样性调查采样点位Table 1 Sampling locations of terrestrial biodiversity survey
1.2.3 物种多样性的测度
研究物种多样性的测度主要采用Margalef丰富度(R)、Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(J)、Sorensen相似性系数(C)等指标。
各指标计算方法及公式分述如下:
(1)Margalef丰富度指数(R):

其中,S为调查样带中的物种数目;N为总个体数。
(2)Shannon-Wiener多样性指数(H):
其中,Pi为第i种的个体数占样本中总个体数的比,如样本总个体数为N,第i种个体数为ni,则Pi=ni/N。
(3)Simpson优势度指数(D):
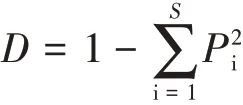
其中,S为物种丰富度指数,Pi为第i种的个体数占样本中总个体数的比,Pi=ni/N。
(4)Pielou均匀度指数(J):
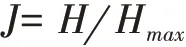
其中,Hmax为最大的物种多样性指数,Hmax=lnS。
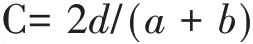
(5)Sorensen相似性系数(C):其中,a和b分别是各样地的物种数,d为两个样地共有的物种数。
2 结果与分析
2.1 植物区系的统计分析
2.1.1 陆地植物的物种组成
通过对8个样点的样地调查统计,调查植物224种,隶属于3门6纲65目71科154属。其中蕨类植物门2纲2目7科7属;被子植物门4纲63目64科147属。其中,被子植物门植物种类最多,为213种,占总数的95.09%;蕨类植物门11种,占总数的4.91%。
(1)蕨类植物大小科的分析
科的大小按属数多少统计,见表2。在花溪陆地8个样点内7科蕨类植物中,各科属含量没有差异,均为1科1属,各占比为100%。相应的科有蕨科(Pteridiaceae)1属、凤尾蕨科(Pteridaceae)1属、姬蕨科(Dennstaedtiaceae)1属、鳞毛蕨科(Dryopteridace⁃ae)1属、水龙骨科(Polypodiaceae)1属、肿足蕨科(Hypodematiaceae)1属、木贼科(Equisetaceae)1属。

表2 蕨类植物科内属组成Table 2 Genus composition within the fern family
科的大小按种数多少统计,见表3。在花溪陆地8个样点内7科蕨类植物中,含2种以上物种有3科,即凤尾蕨科3种、鳞毛蕨科2种、木贼科2种,占总种数的63.64%。含1种的有4科,蕨科1种、姬蕨科1种、水龙骨科1种、肿足蕨科1种,占总种数的36.36%。这些种类在群落中虽然不占主导地位,但是体现了本区植物区系的多样性和复杂性。

表3 蕨类植物科内种组成Table 3 Species composition within the fern family
(2)蕨类植物属的大小分析
属的大小按种数多少统计,见表4。在花溪样点内蕨类植物的7个属中,含2种以上的属有3个,占总属数的42.86%,其中含种数为7种,占总种数的63.64%。含1个种的属有4个,占总属数的57.14%,种数为4种,占总种数的36.36%。由此可见,蕨类植物的种数较为分散且植物成分较少。

表4 蕨类植物属内种组成Table 4 Speciescomposition within the genus Pteridium
(3)被子植物大小科的分析
科的大小按属数的多少统计,见表5。在花溪陆地植被调查内的64科被子植物中,各科含属数差异较大,包含5属以上的科数有4科,占总科数的6.25%,计56属,占比38.1%,在调查内被子植物组成上占重要位置。其中,豆科(Leguminosae)11属,禾本科(Gramineae)13属,菊科(Compositae)25属,蔷薇科(Rosaceae)7属。5属以下的科有60科,占比93.75%,属数91属,占比61.9%。
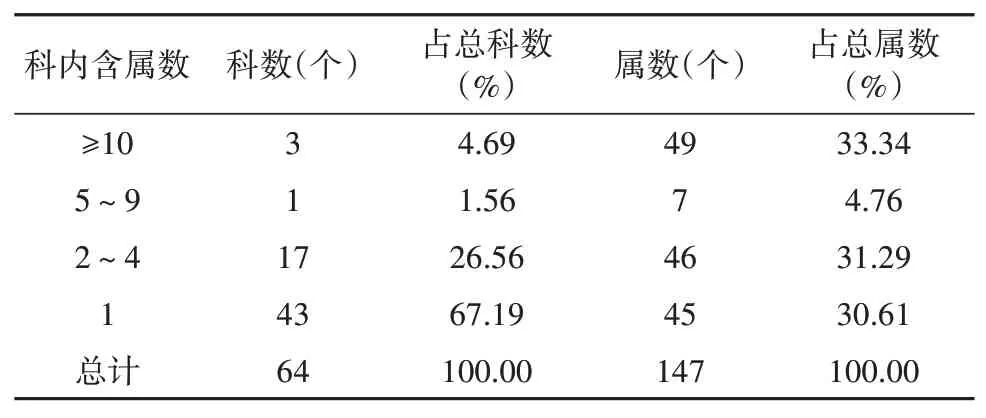
表5 被子植物科内属组成Table 5 Genus composition within angiosperm families
科的大小按种数多少统计(表6)。在花溪陆地植被调查内的64科被子植物中,各科含种数差异较大,包含15种以上的大科有3个,即豆科(11属15种)、禾本科(13属23种)、菊科(25属38种),共计76种,占总种数的35.68%。含5~14科的较大科有5科,占总种数的18.31%,即唇形科(4属7种)、蓼科(4属8种)、葡萄科(4属7种)、蔷薇科(7属12种),桑科(3属5种),含种数有39种,占总种数有18.31%。以上8科占总科数的12.5%,含种数达115种,占总种数的53.99%,在区内占据优势,说明调查区域内优势科明显。含5种以下的小型科和单种科共有56科,占总科数的87.5%,含种数为98种,占总种数为46.01%。这些种类在群落中虽然不占主导地位,但是体现了本区被子植物的多样性和复杂性。
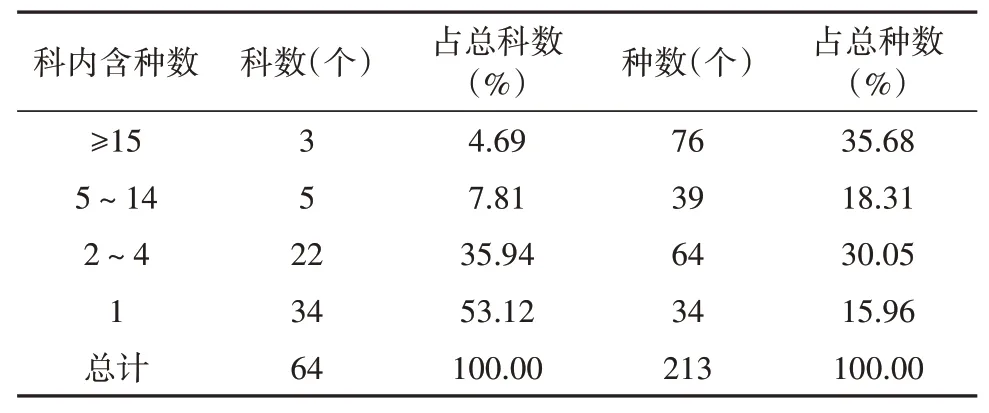
表6 被子植物科内种组成Table 6 Species composition within angiosperm families
(4)被子植物属的大小分析
属的大小按种数多少统计,见表7。在花溪区内调查的147属被子植物中,含5种以上的属有1个,占总属数的0.68%,含5个种,占总种数的2.35%;含2~4种的属共有44属,占总属数的29.93%,种数有106个,占总种数的49.76%;含1个的属有102个,占总属数的69.39%,种数为102个,占总种数的47.89%。由此可见,区内属的组成较为分散,植物成分较为复杂。
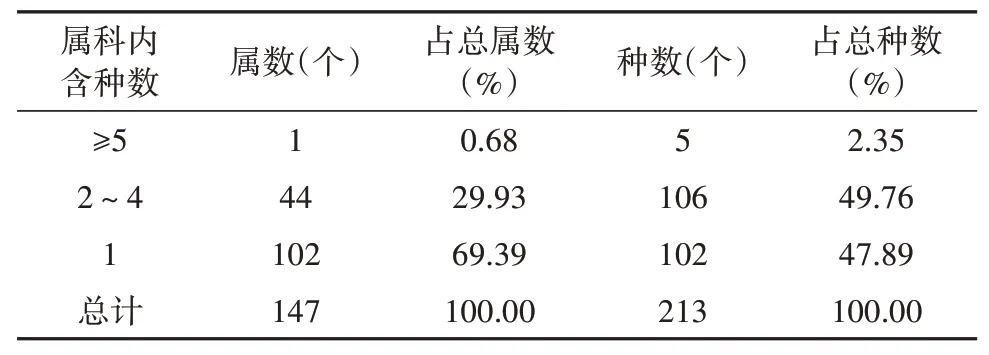
表7 被子植物属内种组成Table 7 Species composition within angiosperm genera
2.1.2 被子植物区系地理成分的统计分析
(1)科的地理成分统计与分析
依据吴征镒等(2003)的《世界种子植物科的分布区类型系统》进行划分,花溪区内被子植物64科分为了11种分布区类型,见表8,可归并为世界分布科、热带分布科、温带分布科。

表8 被子植物科的分布区类型Table 8 Types of distribution areas of angiospermfamilies
1)世界分布科世界广布科共计24科,占总科数的37.5%。如唇形科、豆科、禾本科、菊科、蓼科、葡萄科、蔷薇科、桑科等均是含5种以上的植物科,世界广布科共包含71属115种,分别占总属数45.8%和总种数50.7%。
2)热带分布科热带分布科最多,共有28科,占总科数的43.75%。其中,绝大多数是泛热带分布科,共计18科,含4种以上的科有5个:大戟科(Eu⁃phorbiaceae)、萝藦科(Asclepiadaceae)、葡萄科(Vita⁃ceae)、卫矛科(Celastraceae)、荨麻科(Urticaceae),占总属数的11.56%和总种数的10.8%。
3)温带分布科温带分布科共计12科,占总科数的18.75%,其中北温带及北温带和南温带间断分布类型最多,共有10科。
从科级水平分析花溪样地区内被子植物,世界广布的科数占有明显优势。由表可知,组成花溪内被子植物科的分布区类型比较集中,主要为世界广布和泛热带分布。
(2)属的地理成分统计与分析
依据吴征镒关于中国种子植物属分布区类型的划分,可将花溪内被子植物的147个属划分为13种分布区类型(见表9)。
世界分布的属有19个,占总属数的12.92%,包括飞蓬属(Erigeron)、鬼针草属(Bidens)、千里光属(Senecio)、酸模属(Rumex)、悬钩子属(Rubus)等。
热带分布的属共有65个,占总属数的44.22%,其中泛热带分布的属有31个,占热带分布的47.69%,包括白酒草属(Conyza)、大戟属(Eu⁃phorbia)、牵牛属(Pharbitis)、狼尾草属(Pennise⁃tum)、泽兰属(Eupatorium)等。旧世界热带分布有9个,占热带成分的13.85%。包括艾纳香属(Blumea)、八角枫属(Alangium)、芭蕉属(Musa)等。热带亚洲分布的属有6个,占区内热带成分的9.23%,包括构属(Broussonetia)、山茶属(Camel⁃lia)、五列木属(Pentaphylax)等。热带亚洲和热带美洲间断分布的有4个,占热带成分的6.15%,包括美人蕉属(Canna)、番茄属(Lycopersicon)等。热带亚洲至热带大洋洲分布的属有5个,占热带成分的7.69%,包括姜属(Zingiber)、水麻属(Debrege⁃asia)等。热带亚洲至热带非洲分布的属有10个,占热带成分的15.39%,包括姜花属(Hedychium)、荩草属(Duthiea)、菊三七属(Gynura)、芒属(Mis⁃canthus)等。
温带分布的属共63个,占总属数的42.86%,与热带分布属相差不大。其中北温带分布属有25个,占温带成分的39.68%,包括百合属(Lilium)、蔷薇属(Rosa)、蒿属(Artemisia)、何首乌属(Fallopia)、车轴草属(Trifolium)等,是温带分布属最主要的成分。旧世界温带分布的属有10个,占温带分布属的15.87%,包括菊属(Chrysanthemum)、女贞属(Li⁃gustrum)、荞麦属(Fagopyrum)等。温带亚洲分布的属只有1个,马兰属(Kalimeris),占温带成分的1.59%。东亚分布的有17个,占温带成分的26.98%,包括半夏属(Pinellia)、败酱属(Patrinia)、刺楸属(Kalopanax)、刚竹属(Phyllostachys)等。东亚和北美洲间断分布的属有9个,占温带成分的14.29%,包括地锦属(Parthenocissus)、胡枝子属(Lespedeza)、漆属(Toxicodendron)等。中国特有分布的属有1个,银杏属(Ginkgo),占温带成分的1.59%。
从表9可以看出,花溪样地内被子植物有13个分布类型,说明本区被子植物区系地理成分复杂,分布区类型多样。最多的为泛热带分布,占总属数的21.09%,其次为北温带分布,占总属数的17.01%。世界分布与东亚分布相差不大,分别为19属(占总属数12.93%)和17属(11.57%),这两个分布类型也是广域性分布,这4个广域性分布型共92属,占总属数的62.59%,表明区域内被子植物具有隐域性特征。从表10可知,被子植物热带成分分布属有65个,温带成分63个,热带类型略多于温带,说明区内被子植物具有亚热带和暖冬带的双重性质。
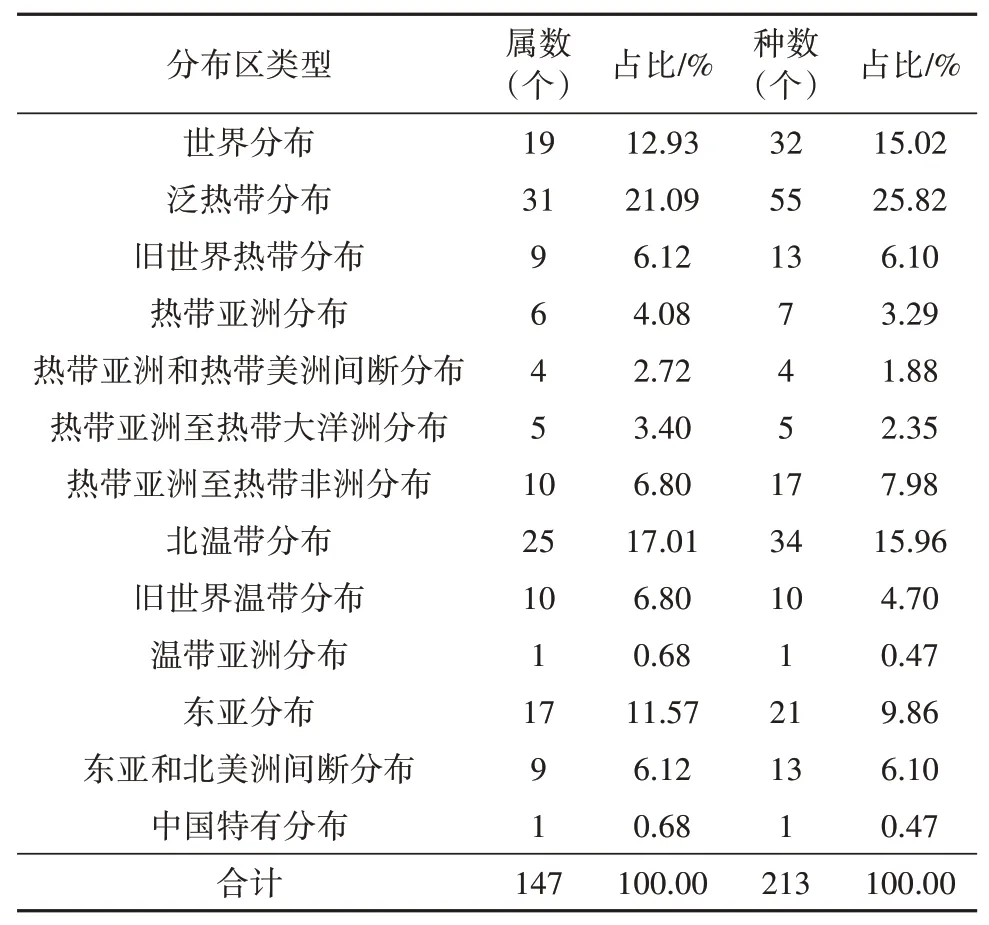
表9 花溪样地内被子植物属的分布区类型Table 9 Distribution area types of angiosperm genera in the Huaxisample site
2.1.3 蕨类植物分布区类型分析
(1)科的分布区类型分析
表10 中显示,在花溪植被调查中,蕨类植物的科的只有3个分布区类型,各分布类型占比不同,世界广布占比57.14%,分别是蕨科、鳞毛蕨科、水龙骨科、木贼科,泛热带分布是凤尾蕨科、姬蕨科,占总科数的28.57%,热带亚洲至热带非洲分布是肿足蕨科,占总科数的14.29%。

表10 蕨类植物科的分布区类型Table 10 Types of distribution areas of fern families
(2)属的分布区类型分析
蕨类植物的属的有7个分布区类型,各分布类型分别占一个属,占比均是14.29%,世界分布是蕨属,泛热带分布是凤尾蕨属,泛亚热带分布是姬蕨属,温带亚洲分布是贯众属,热带亚洲分布是石韦属,热带亚洲至热带非洲分布是肿足蕨属,北温带分布是木贼属。
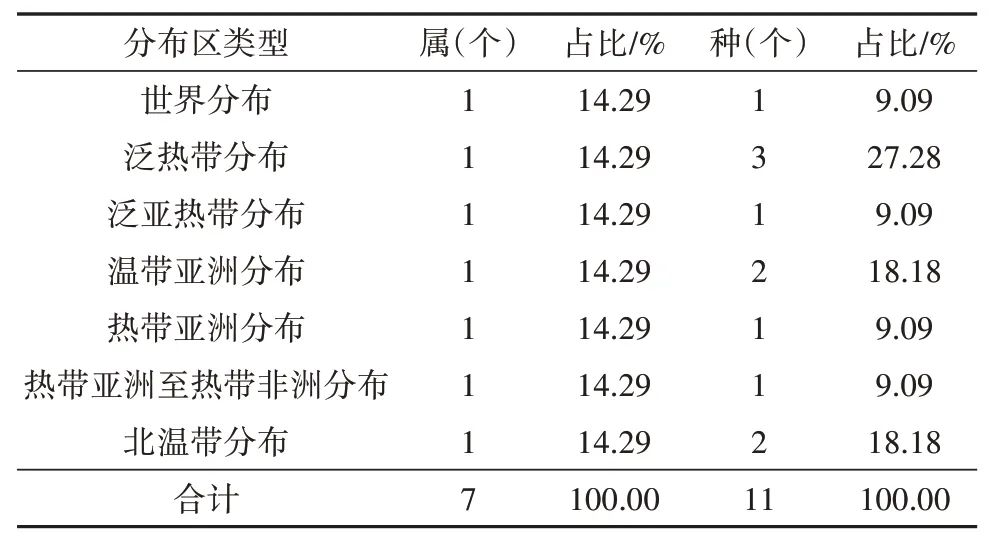
表11 蕨类植物属、种的分布区类型Table 11 Types of distribution areas of fern genera and species
2.2 植物群落物种多样性
2.2.1 植物α多样性变化
植物多样性的各个指数都反映了相应的群落及生境情况。Shannon-Wiener指数和Simpson指数可以较好的反应群落的结构,而均匀度是指一个生境或群落中个体数目的分配情况,反映了各物种组成的均匀程度[15]。
在本次植物多样性调查中,每个样地作为一个整体研究,对样地之间的多样性进行比对分析,寻找样地间存在差异的原因。结果表明:Pielou均匀度指数和Simpson指数表现总趋势基本一致,Margalef指数与Shannon-Wiener指数呈现下降后上升趋势,主要是由于在同一海拔梯度,越靠近水库位置的物种丰富度较高。样地Ⅰ靠近花溪水库,而样地Ⅵ、样地Ⅶ和样地Ⅷ靠近阿哈水库,随着与水库距离发生变化,物种丰富度也呈现变化趋势,8个样地的中间段也受公路和人为因素的影响,呈现下降趋势,也由于各样地草本层物种数量和种类存在差异。各样地丰富度指数依次是样地Ⅶ>样地Ⅷ>样地Ⅰ>样地Ⅵ>样地Ⅲ>样地Ⅴ>样地Ⅳ>样地Ⅱ,Shannon-Wiener指数依次是样地Ⅷ>样地Ⅰ>样地Ⅶ>样地Ⅵ>样地Ⅲ>样地Ⅴ>样地Ⅳ>样地Ⅱ,Pielou均匀度指数依次为样地Ⅳ>样地Ⅲ>样地Ⅱ>样地Ⅰ>样地Ⅴ>样地Ⅷ>样地Ⅵ>样地Ⅶ,Simpson指数依次为样地Ⅰ>样地Ⅷ>样地Ⅲ>样地Ⅵ>样地Ⅳ>样地Ⅶ>样地Ⅱ>样地Ⅴ,见图1。
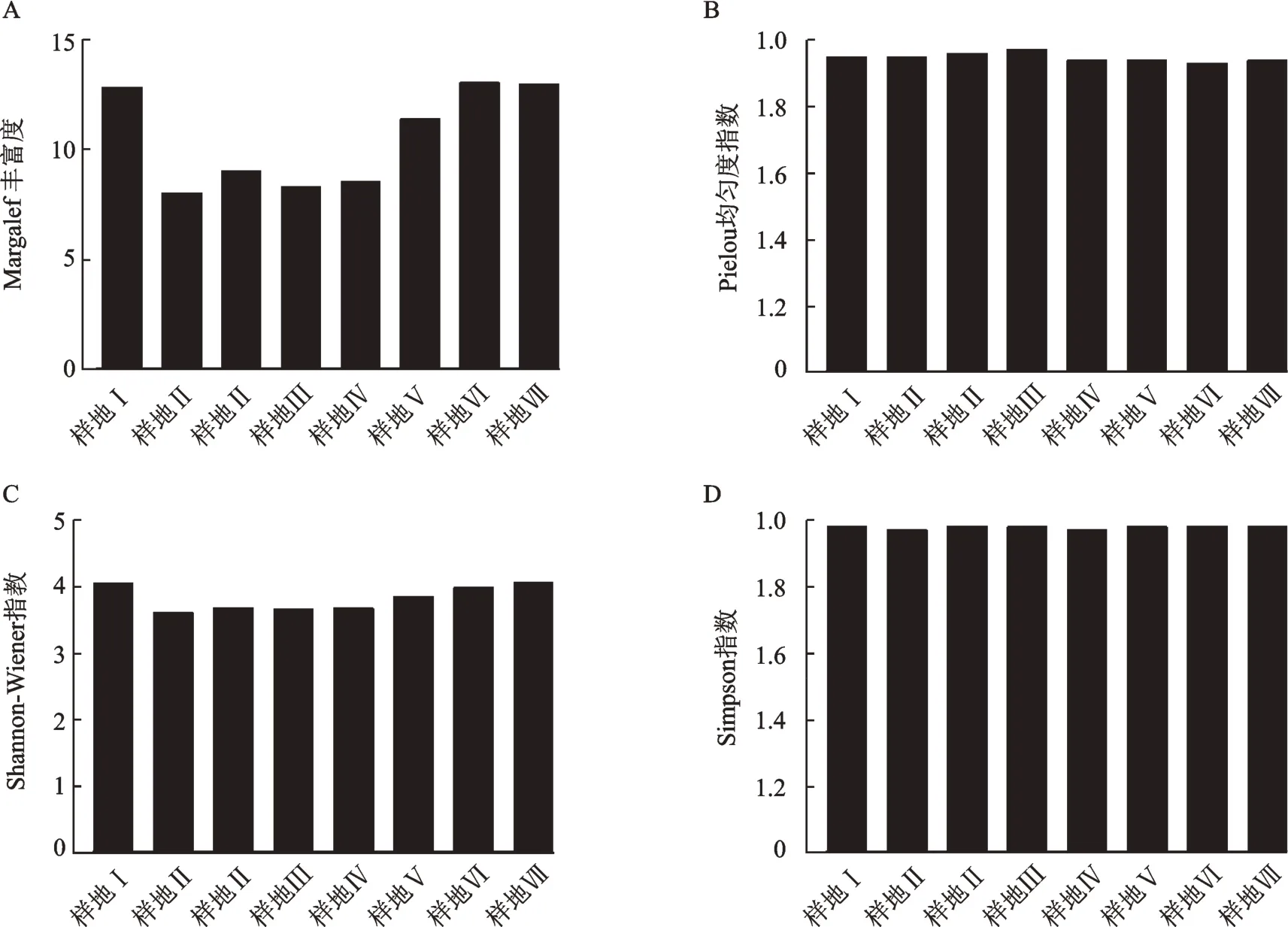
图1 样地植物多样性指数Fig.1 Plant diversity indexes of sample sites
2.2.2 Sorensen相似性系数
从Sorensen相似性系数(表12)中可以看出,最高为样地Ⅲ和样地Ⅳ,样地Ⅵ和样地Ⅷ,都达到0.53,其次为样地Ⅳ和样地Ⅴ,系数为0.52,样地Ⅱ和样地Ⅲ,为0.47。样地Ⅰ和样地Ⅴ最低,相似性系数为0.18。这基本说明样地相邻越近植物多样性越相似,植物共有数也越多。

表12 Sorensen相关性系数Table 12 Sorensen correlation coefficients
3 结论与讨论
通过对花溪区花溪水库至阿哈湖水库的8个样地进行植物群落物种组成及多样性进行研究,结果表明花溪水库物种多样性相对较高,共有被子植物64科147属213种,蕨类植物7科7属11种。被子植物区系地理成分复杂,分布区类型多样,科的分布区类型以世界广布占明显优势(37.5%);属的分布区类型以热带成分占主要优势(44.21%),其次是温带成分分布(42.86%),该区植物表现出了突出的复杂性和差异性。物种组成最为丰富的是样地Ⅶ,样地Ⅱ、样地Ⅳ、样地Ⅴ则种类较少,这主要由于人为活动频繁,对环境影响较大,交通枢纽及一些工程建设场地占用,导致物种适应能力较弱,无法形成更为稳定的植物群落,从而造成生物多样性降低、植物类型减少。
在α多样性的调查中我们可以得知,物种的丰富度分布各一,这是自然因素和人为因素综合作用下的结果,使植物群落分布各有不同[16]。在靠近花溪水库的样地Ⅰ和靠近阿哈湖水库的样地Ⅶ、样地Ⅷ的α多样性都处于较高优势水平,可能是因为靠近水源的地方,水资源相对丰富,能够充分提供植物生长的水分需求。Margalef指数显著高于其他样地,Shannon-Wiener指数也呈现高于其他样地的趋势,Pielou均匀度指数和Simpson指数整体较为一致,没有明显变化。样地多样性体现所处的地理环境和自然资源有着明显的关系,各种环境因子都影响着植物的生长及多样性的变化,而人为因素及公路建设也影响着多样性的高低。同时,外来物种的入侵及化感作用,也使入侵区域的植物多样性降低[17],样地内苍耳、茅草等中生、旱生植物逐渐替代,逐渐成为样地内优势种,某些湿生植物群落渐变为旱生植物植物群落[18],而随着旱生植物的增加,对原有的湿地环境产生影响[19]。
实地调查发现,在样地区域周围,附近施工工程也影响着周围的自然环境,施工产生的废弃物对周围植被的生长也有影响,且人类活动较为频繁,烧荒和开垦导致生境退化,乔本植物在部分区域数量较少,草本植物种类单一,且部分区域外来物种入侵也比较严重。因此,保护和监测饮用水源地周围的植被是至关重要,保留更为原生态的自然环境,为人类提供干净卫生的水资源,为动植物提供更好的生存环境,这更有利于人们的生产生活,对施工区域进行施工后的植被资源恢复,也是必不可少的。因此,加强饮用水源地保护区周围环境的保护治理,退耕还湿,严格控制入侵植物对生态环境的破坏,同时需要加强宣传力度和提高居民保护意识,造就绿水青山的环境,人与自然和谐共生的景观。
猜你喜欢
福建林业科技(2023年4期)2024-01-02 04:06:02
安徽农学通报(2021年22期)2021-12-11 20:37:03
安徽农业科学(2021年14期)2021-09-03 09:55:59
乡村地理(2021年4期)2021-03-28 06:26:55
耕作与栽培(2020年3期)2020-09-18 09:13:00
文苑(2020年5期)2020-06-16 03:18:40
花卉(2020年12期)2020-01-09 07:12:52
炎黄地理(2019年5期)2019-10-20 01:59:19
乡村地理(2017年4期)2017-09-18 02:53:54
安徽农业科学(2016年9期)2016-10-14 22:12:23
