
30个野生灵芝菌株遗传多样性及农艺性状评价
2022-02-15 01:02张彬彬王震刘保卫赵翠敏郭金英王春霞郑素月
江苏农业科学 2022年2期
张彬彬 王震 刘保卫 赵翠敏 郭金英 王春霞 郑素月





摘要:为了给灵芝的生产、驯化及遗传育种提供种质资源,采用酯酶同工酶技术、分子标记技术(ISSR),结合出菇形态学分析了30个野生灵芝菌株的遗传多样性及农艺性状,构建了聚类树状图。酯酶同工酶分析结果表明,当相似系数在0.709时,可以将30个野生灵芝菌株分为7类;ISSR分析结果表明,当相似系数在0.728时,可以将30个菌株分为6类。出菇试验结果表明,30个菌株中有25个菌株能出菇,按形态特征,在欧氏距离为3.92时可将25个灵芝菌株分为五大类群,3种方法存在一致性。通过驯化栽培及农艺性状比较分析筛选出5个产量较高、品相较好的菌株。30个野生灵芝菌株具有较为丰富的遗传多样性。
关键词:野生灵芝;酯酶同工酶;ISSR;农艺性状;多样性
中图分类号:S567.3+10.2 文献标志码: A
文章编号:1002-1302(2022)02-0108-06
收稿日期:2021-05-06
基金项目:河北省现代农业产业技术体系食用菌创新团队建设专项(编号:HBCT2018050202)。
作者简介:张彬彬(1988—),女,河北保定人,硕士研究生,主要从事食用菌种质资源研究。E-mail:921940953@qq.com。
通信作者:郑素月,博士,教授,硕士生导师,主要从事食用菌种质资源研究。E-mail:zhengsuyue@hebeu.edu.cn。
灵芝是我国传统的中药材,深受消费者喜爱[1]。现代研究表明,灵芝含有多种活性成分,如灵芝多糖、三萜、麦角甾醇、生物碱等[2],在抗癌、防癌、抗糖尿病、提高免疫力和抗氧化等方面有极高的药用价值[3-7]。灵芝属于中高温栽培种类,野生资源广泛分布于东亚的暖温带和亚热带地区,在我国自然分布于河北、山东、河南、安徽、江苏、浙江、江西、湖北、湖南、四川和云南等地[8]。长期以来,可供人工栽培和加工利用的灵芝品种比较单一,不能满足多样化的市场需求。因此,野生灵芝种质资源的开发利用及其人工驯化研究具有重要意义。酯酶同工酶技术和分子标记技术(ISSR)检测手段简单、高效,广泛应用于食用菌的菌种选育、分类鉴定和种质资源评价等研究[9-10]。灵芝属的多数种类生长在阔叶树的腐木上,河南省南召县森林覆盖率达66.49%,为灵芝提供了良好的生存环境。目前,笔者未见国内外文献对南召县灵芝资源的开发利用和系统深入地研究。因此,本试验对河南省南召县的野生灵芝进行调查收集,通过生化、分子标记技术和形态学鉴定3种方法对其亲缘关系进行初步的研究,通过人工驯化筛选优良品种,以期为灵芝生产栽培和遗传育种提供种质资源。
1 材料和方法
1.1 试验材料
1.1.1 供试菌株
供试野生灵芝菌株30个,均于2018年8月从河南省南召县采集,由河北工程大学食用菌实验室分离和保藏,菌株编号见表1。
1.1.2 供试培养基
(1)马铃薯葡萄糖琼脂(PDA)培养基。
(2)木屑培养基:木屑(煮汁) 200 g、豆粕(煮汁)30 g、葡萄糖20 g、琼脂粉20 g、蛋白胨4 g、硫酸镁1 g、磷酸二氢钾3 g。
(3)原种培养基:小麦95.5%、石灰2%、石膏1%、磷酸二氢钾1%、硫酸镁0.5%。
(4)栽培种培养基:棉籽壳86%、麦麸12%、石灰1%、石膏1%。
1.2 试验方法
试验于2019年11月至2020年12月在河北工程大学食用菌实验室完成。酯酶同工酶分析采用垂直板聚丙烯酰胺活性凝胶电泳方法[11]。ISSR分子引物从28个常用引物[均由生工生物工程(上海)有限公司合成]中筛选合适的引物,对30个灵芝菌株进行聚合酶链式反应(PCR)扩增,PCR扩增体和电泳条件参照文献[12-13]所述方法。形態学鉴定将活化好的菌种定量(5 mm2)接种到PDA综合培养基上,置于25 ℃恒温培养箱培养,观察菌丝形态,记录菌丝生长性状。出菇形态采用熟料栽培,常规出菇试验,观察子实体形态, 记录子实体形态特征,测量菌盖直径、菌盖厚度、菌柄长度、生物转化率等。
1.3 数据处理
利用NTSYSpc分析软件构建各菌株的亲缘关系树状图。按易腾飞等的方法[14]计算遗传多样性指数(H′),利用DPS 7.05软件进行聚类分析。
2 结果与分析
2.1 酯酶同工酶结果分析
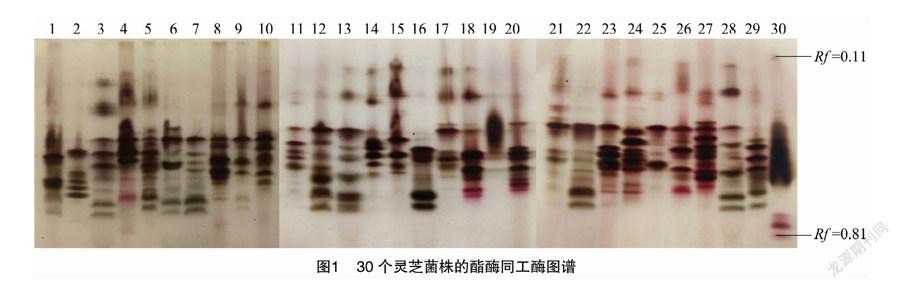
如图1所示,菌株的酶谱迁移率(Rf)主要介于0.11~0.81之间,30个菌株共扩增出30条不同酶带,多态性较高。对其电泳图谱进行相似性聚类分析,由图2可知,30个灵芝菌株两两间相似系数的分布范围是0.65~0.93,当相似系数在0.709时,可以将30个野生灵芝菌株分为7类:第1类包括20个菌株,分别为YL-15、YL-8、YL-42、YL-12、YL-72、NZ-33、YL-53、YL-79、NZ-27-1、YL-37、YL-82、YL-77、YL-68、YL-83、YL-49、YL-51、YL-40、YL-31、YL-61;第2类为 YL-18;第三类为YL-90、YL-65;第4类为YL-57和YL-93;第5类为YL-88和YL-89;第6类为 NZ-28 和YL-76;第7类为YL-81和YL-14。
2.2 30个野生灵芝菌株ISSR分子鉴定结果与分析
通过对28个常用引物进行ISSR分析,筛选出7个对灵芝菌株DNA扩增重复性好、条带清晰且有明显多态性片段的引物,引物序列见表2。
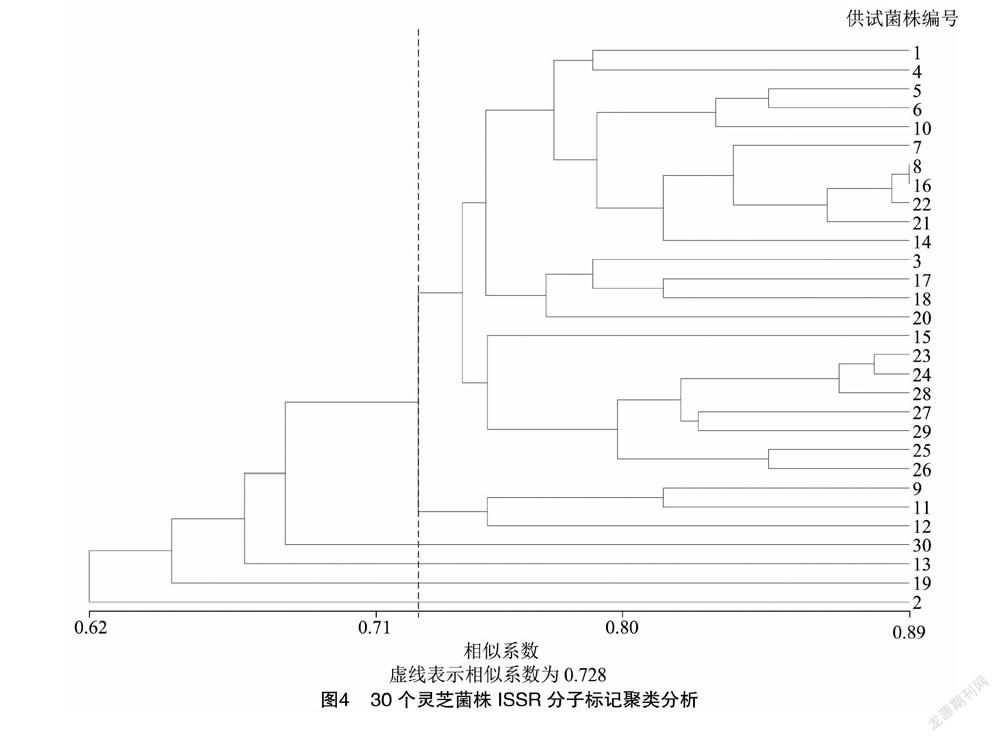
用这7个引物对30个灵芝菌株的基因组进行扩增,可获得多态性强、稳定性好且具有一定特异性的DNA指纹图谱,部分引物扩增结果见图3。经统计,7个引物共扩增出85条清晰的多态性片段,DNA片段的大小介于300~8 000 bp之间,平均每个引物对野生灵芝菌株基因组扩增的条带为12.1条。将30个野生灵芝菌株的ISSR图谱转换为数字矩阵,用NTSYS软件进行聚类分析,得到供试菌株的亲缘关系树状图(图4),30个灵芝菌株遗传系数在0.62~0.89之间。相似系数在0.728时,30个菌株可以分为6类:第1类包括23个菌株,分别为YL-15、YL-57、YL-93、YL-42、YL-49、YL-12、YL-68、NZ-33、YL-79、YL-37、YL-90、YL-8、YL-40、YL-88、YL-89、YL-31、NZ-28、YL-76、YL-81、YL-77、YL-14、YL-82和YL-53;第2类为YL-83、YL-51 和YL-61;第3类为YL-18;第4类为YL-72;第5类为NZ-27-1;第6类为YL-65。说明这30个野生灵芝菌株具有丰富的遗传多样性。
2.3 形态学鉴定
2.3.1 菌丝形态
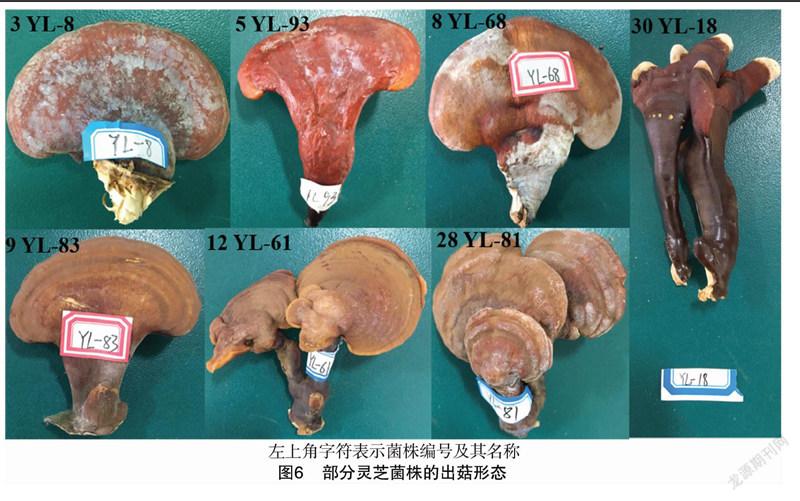
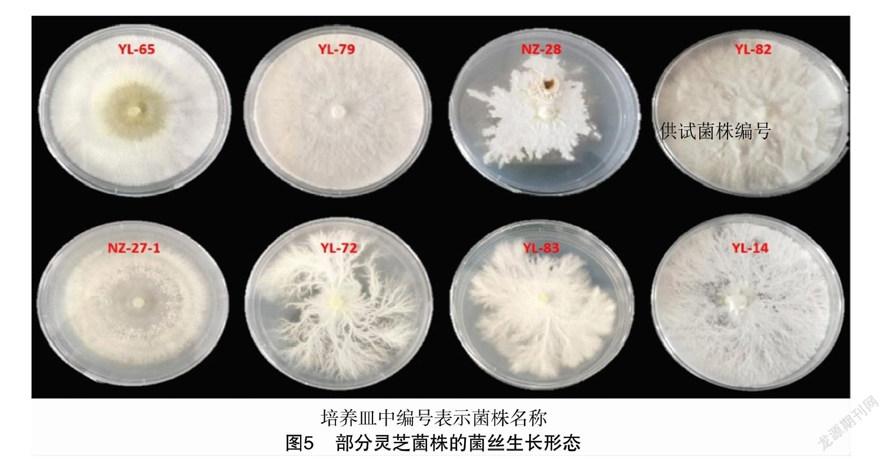
不同灵芝菌株的菌丝形态差异较大,可将30个菌株的形态类型分为四大类,部分代表菌株形态如图5所示,分类结果和描述见表3。
2.3.2 子实体农艺性状
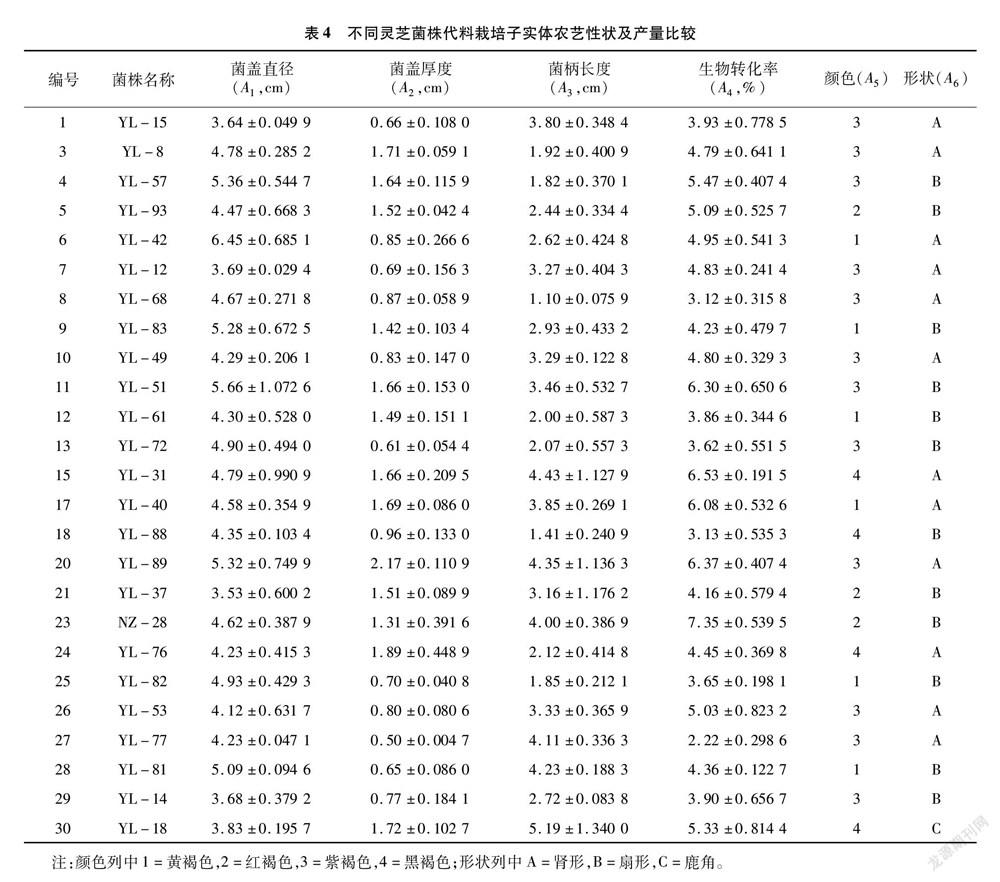
对30个野生灵芝进行驯化栽培,成功得到25个灵芝菌株的子实体,部分子实体形态见图6。对其4个数量性状和2个质量性状进行记录,结果见表4。多样性分析结果(表5)表明,数量性状遗传多样性指数以生物转化率(A4)最高,为2.110 6,其次依次为菌盖直径(A1,1.981 5)、菌柄长度(A3,1.978 7)、菌盖厚度(A2,1.779 5);质量性状中颜色的遗传多样性指数(1.242 5)高于形状的遗传多样性指数(0.833 4)。
如表5所示,不同菌株间变异系数存在很大差异,菌盖厚度的变异系数最大,为40%,变幅为 0.50~2.17 cm;变异系数最小的是菌盖直径,为14.9%,变幅为3.53~6.45 cm。
2.3.3 农艺性状的聚类分析
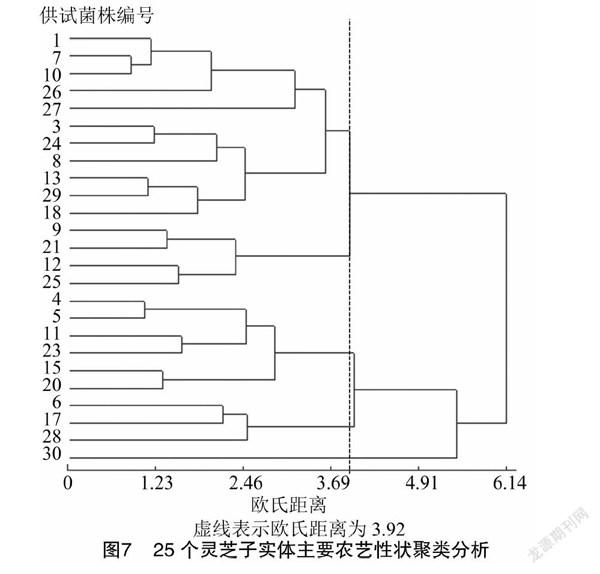
本研究采用最长距离法对不同灵芝菌株的欧氏距离进行聚类分析,绘制聚类树状图。如图7所示,在欧氏距离为3.92时可将25个灵芝菌株分为五大类群:第1类包括11个菌株,这一类多为紫褐色肾形或扇形灵芝,产量较低,生物转化率仅为2.22%~5.03%;第2类有4个菌株,这一类多为黄褐色扇形灵芝;第3类有6个菌株,包括红褐色扇形灵芝,产量最高高,生物转化率最高可到7.35%;第4类包括YL-42、YL-40 和YL-81等3个菌株,为黄褐色灵芝,产量较高;第5类为1株黑褐色的鹿角灵芝。
3 结果与讨论
本研究以南召县30个野生灵芝菌株为供试材料,从生化、ISSR分子标记和形态学3个方面研究了其亲缘关系。菌株的酯酶同工酶谱Rf值主要介于0.11~0.81之间,30个灵芝菌株两两间相似系数的分布范围是0.65~0.93,当相似系数在0.709时,可以将30个野生灵芝菌株分为7类。ISSR筛选出的7个引物共扩增出85条清晰的多态性片段,DNA片段的大小介于300~8 000 bp之间,30个灵芝菌株遗传相似性在0.62~0.89之间,相似系数在0.728时,30个菌株可以分为6类。通过6个主要农艺性状对25个驯化成功的灵芝菌株进行聚类,结果显示,当欧氏距离为3.92时,可将25个灵芝菌株分为五大类群。本研究中,酯酶聚类(图2)、ISSR聚类(图4)和实体形态分类(图7)等3种方法都把YL-15、YL-8、YL-12、YL-68、YL-49、YL-53和YL-77这7个菌株聚为一类,同时也把鹿角灵芝YL-18分为一类。为增加结果的准确性,可以联合使用3种分类方法对野生灵芝菌株亲缘关系进行鉴定。通过驯化栽培及农艺性状比较分析,筛选出5个产量较高、品相较好的菌株NZ-28、YL-31、YL-89、YL-51和YL-18。
本研究有5个菌株未驯化成功,其中NZ-33有原基产生,但是后期未分化成子实体;其余4个菌株YL-65、YL-90、NZ-27-1和YL-79未出现原基。除NZ-27-1外,其余4个菌株菌丝生长性状良好,未出菇原因有待进一步研究。本研究为河南省南召县野生灵芝资源的保护和合理开发利用提供了理论依据,同时也为建立野生灵芝种质资源数据库奠定了基础。
参考文献:
[1]李钦艳,钟莹莹,李 忠,等. 不同生长阶段灵芝子实体和孢子粉活性成分分析[J]. 中国农学通报,2021,37(1):49-54.
[2]张瑞婷,周 涛,宋潇潇,等. 灵芝活性成分及其药理作用的研究进展[J]. 安徽农业科学,2018,46(3):18-19,22.
[3]Bai J H,Xu J,Zhao J,et al. Ganoderma lucidum polysaccharide enzymatic hydrolysate suppresses the growth of human colon cancer cells via inducing apoptosis[J]. Cell Transplantation,2020,29(1):1-9.
[4]徐 锦,汪雯翰,楊 妍,等. 乙醇浓度对提取灵芝三萜含量的影响及提取物抗前列腺癌细胞LNCaP的活性[J]. 菌物学报,2020,39(1):155-163.
[5]Chiu C H,Wang R,Zhuang S,et al. Biotransformation of mogrosides from Siraitia grosvenorii by Ganoderma lucidum mycelium and the purification of mogroside Ⅲ E by macroporous resins[J]. Journal of Food and Drug Analysis,2019,28(1):74-83.
[6]Liu H Z,Qiu F F,Wang Y Y,et al. A recombinant protein rLZ-8,originally extracted from Ganoderma lucidum,ameliorates OVA-inducedlunginflammationbyregulatingTh17/Tregbalance[J]. Journal of Leukocyte Biology, 2020,108(2):531-545.
[7]田淑雨,鹿士峰,吴杨洋,等. 超声破碎辅助提取灵芝多糖工艺优化及抗氧化活性研究[J]. 食品研究与开发,2019,40(8):101-107.
[8]戴玉成,曹 云,周丽伟,等. 中国灵芝学名之管见[J]. 菌物学报,2013,32(6):947-952.
[9]孔祥会,姚方杰,王 鹏. 食用菌种质资源评价方法及在品种选育上的应用实践[J]. 中国食用菌,2019,38(12):8-10.
[10]孟 虎,孙国琴,睢 韡,等. ISSR技术在食用菌研究上的应用[J]. 北方园艺,2016(5):207-210.
[11]郭金英,宋彦龙,李 超,等. 四十个野生香菇菌株遗传多样性分析[J]. 北方园艺,2018(10):157-160.
[12]李辉平. ISSR在食用菌遗传多样性研究中的应用[D]. 北京:中国农业科学院,2007:26-27
[13]中华人民共和国农业部.食用菌菌种真实性鉴定 ISSR法:NY/T 1730—2009[S].北京:中华人民共和国农业部,2009.
[14]易腾飞,李珊珊,李嘉豪,等. 261份小麦品种基于农艺性状的遗传多样性分析[J]. 河北农业大学学报,2018,41(2):7-13.
猜你喜欢
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
求知导刊(2016年30期)2016-12-03
戏剧之家(2016年22期)2016-11-30
中国中药杂志(2016年20期)2016-11-19
科技资讯(2016年19期)2016-11-15
人间(2016年27期)2016-11-11
农业与技术(2016年15期)2016-11-09
