
抚育间伐对小兴安岭天然次生林中杉木的碳密度分配与竞争的影响
2022-02-10 12:35王子纯李耀翔孟永斌
西北林学院学报 2022年1期
王子纯,李耀翔,孟永斌,王 晨
(东北林业大学,黑龙江 哈尔滨 150040)
抚育间伐是以促进森林生长、提高林木质量为目的,是一种以培育目标树,对林木进行伐密留疏、伐坏留好的作业。抚育间伐必然影响森林种群的结构,从而影响林分生物量、碳密度及竞争关系[1-3]。天然次生林是中国森林的重要组成部分,对维护生态平衡有着较大的贡献,在林分环境、结构组成和生产力等方面与人工林和原始林显著不同[4],目前就天然林分别在土壤性质[5]、水文功能[6]、林下植被[7]等方面展开了较多研究,也有学者对间伐后林分碳密度变化展开研究,发现林分碳密度与树种、培育措施、间伐强度和间伐后持续时间等多种因素有关[8]。本研究以小兴安岭带岭林业局东方红林场天然针阔混交次生林为对象,对不同抚育间伐强度林分进行径阶分布、生物量、碳密度及竞争指数的研究,为小兴安岭林区碳库管理及科学经营提供参考。
1 研究区概况
研究区位于小兴安岭地区伊春市带岭林业局东方红林场414林班(129°5.150′-129°5.205′E,46°52.511′-46°52.827′N)。地处中温带,属于大陆性湿润季风气候,夏季短且温热湿润,冬季长且干燥寒冷、多雪,全年平均气温1.4 ℃。降水主要集中在7-8月,年平均降水量为661 mm。试验样地平均海拔高度为493.36 m,位于山的中腹。研究区林分类型为针阔混交天然次生林,主要树种有冷杉(Abiesfabri)、色木槭(Acermono)、云杉(Piceaasperata)、青楷槭(Acertegmentosum)、水曲柳(Fraxinusmandschurica)、黄波椤(Phellodendronamurense)、红松(Pinuskoraiensis)、紫椴(Tiliaamurensis)、杨树(Populus)、山桃(Prunusdavidiana)、花楷槭(Acerukurunduense)等。
2 研究方法
2.1 研究样地的布设与调查
天然林试验区于2012年进行带状抚育间伐,选取小兴安岭天然林试验区内3个抚育间伐强度,分别近似为15%(样地编号T15)、25%(样地编号T25)、35%(样地编号T35)(以林分密度计),应用下层抚育法,采伐濒死木、被压木及干型不良的林木,同时在间伐区进行补植作业,补植树种包括红松、云杉、落叶松(Larixgmelinii)[9]。2019年8月进行样地复查,在3种间伐强度的样地内分别设置20 m×20 m的标准试验样地各3块,在保留带未间伐林分选取与间伐林分条件相似的林分设置3块20 m×20 m标准试验样地作为对照样地(CK),共计12块样地。间伐林分基本状况见表1。

表1 间伐林分基本状况
2.2 径阶分布检验方法
采用“偏度-峰度系数+Shapiro-Wilk检验”的方法,对大小兴安岭试验样地杉木径阶正态特征进行分析。
2.2.1 偏度、峰度系数的计算 偏度是描述数据分布形态的统计量(在公式中用SK表示),是某总体取值分布的对称性的特征统计量。峰度与峰度类似表征概率密度分布曲线在平均值处峰值高低的特征数(在公式中用KT表示),本研究用来表示林木径阶分布特征。
(1)
(2)
式中:SK表示偏度;KT表示峰度;n为样本个数;xi为样第i个样本值;x为样本平均值;s为样本的标准差。
偏度的绝对值数值越大表示林木径阶分布形态的偏斜程度越大,峰度的绝对值数值越大表示林木径阶分布形态的陡缓程度与正态分布的差异程度越大。
2.2.2 Shapiro-Wilk检验P值 利用Shapiro-Wilk检验(S-W检验)对各林层径级分布进行正态性检验。原假设H0为总体服从正态分布,备择假设H1为总体不服从正态分布。利用S-W检验原假设H0的过程如下。
先将检验样本值升序排列:
x1≤x2≤…≤xn
(3)
计算S-W检验的统计量W:
(4)
在95%的置信区间内,如果W≥0.05,则表示接受原假设,服从正态分布;否则,拒绝假设,认为总体不服从正态分布。
2.3 竞争指数
Hegyi把竞争木株树n确定在半径3.05 m内的所有树木[11],每个样地选取6株对象木,用所有对象木的平均竞争指数代表该样地林木竞争水平[12]。
竞争指数:
(5)
式中:CIi为林木i的竞争指数;Lij为目标树i与竞争树j之间的距离;di、dj为目标树i与竞争树j的胸径;n为竞争树木的株数。
平均竞争指数CI(在公式中用CI表示)。
(6)
式中:CI为林分竞争指数;CIi为林木i的点竞争指数;N为林木总株数;
2.4 林分碳密度计算方法
利用东北林区现有的林木生物量方程,结合每木检尺调查数据,计算出各标准试验样地内优势树种的生物量。云杉和冷杉生物量模型采用胡海清等[13]建立的小兴安岭典型林分生物量模型。样品含碳率的测定采用重铬酸钾氧化-外加热法。
单木各器官生物量乘以该树种在相应器官的含碳率合计即为单木碳储量[14],累积一个样地的单木碳储量则为该样地总碳储量,单位面积的总碳储量则为碳密度(t·hm-2)[15]。
(7)
2.5 数据处理
采用Excel 2010和Origin85进行数据的整理、分析以及图表的绘制,用SPSS 17.0进行单因素方差分析(one-way ANOVA)及Duncan多重比较差异显著性检验(α=0.05),冗余分析(RDA)采用Canoco5软件进行。
3 结果与分析
3.1 抚育间伐强度对林木的径级分布特征的影响
以4 cm作为径阶对杉木大小多样性进行研究,并根据实测数据将杉木划分为3个径级,分别对应杉木的小径级,中径级和大径级3个阶段,具体径级划分为Ⅰ级:5 cm≤DBH<16 cm,Ⅱ级:16 cm≤DBH<26 cm,Ⅲ级:DBH≥26 cm。间伐改变了杉木的径阶分布比例(图1),与CK相比,T15多出6.47%的小径级杉木,说明林分更新效果好,但大径级杉木所占比例最少,仅占总数的16.13%;而T25样地中,中径级和大径级林木所占比例与CK相似,共占65.52%;T35样地中径阶为22 cm的杉木消失,大径阶杉木明显增多,占总数的33.48%,说明该样地杉木生长良好。
对杉木进行径阶正态性检验,由杉木的径阶正态性检验表(表2)可以看出,在T15、T25和T35样地中,杉木的径阶结构较对照样地有明显差异,间伐处理使峰度由负变正。杉木径阶正态性检验结果表明,较大的间伐强度使径阶分布曲线偏离正态性(P<0.05)。
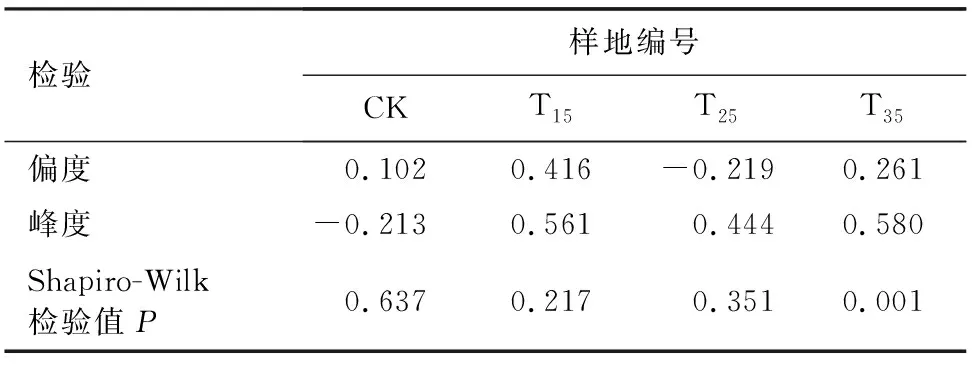
表2 林木的径阶正态性检验
3.2 抚育间伐强度对杉木生物量的影响
由表3可知,间伐强度为15%、25%、35%时,杉木的地上部分生物量均高于对照组,并且随着间伐强度增大呈先增加后减少的趋势,在间伐强度为25%时最大,为对照样地的1.31倍,由此可见,小兴安岭天然次生林间伐8 a后,25%间伐强度最有利于杉木恢复。
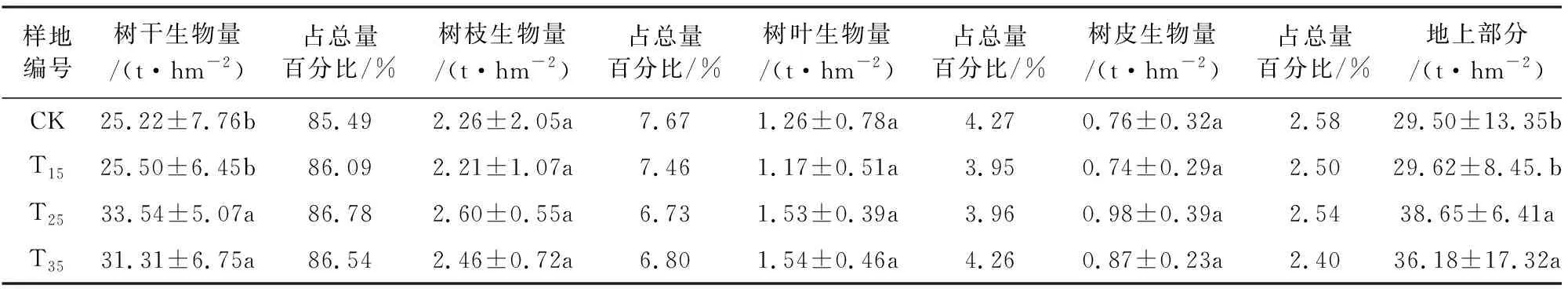
表3 不同间伐强度下林分生物量(平均值±标准差)
树干生物量占总生物量比例最大,为85.49%~86.78%,随间伐强度增加树干生物量所占比例呈先增加再减少的趋势,在T25样地树干生物量占总量百分比最大,这是由于间伐强度较小时林分密度较大,较多的林木争夺有限的资源,不利于树干生物量累积。间伐强度较大时,水源涵养功能减弱,土壤养分元素含量下降,对树干生物量增长也是不利的,因此树干生物量占总量百分比减少,而T25样地环境因子综合作用利于树干生物量增长,因此占总量百分比最大。
树枝生物量占总量的6.73%~7.67%,CK样地树枝生物量为2.26 t·hm-2,占总量百分比最大,为7.67%,而间伐整体使树枝生物量占比减小,当间伐强度为15%、25%和35%时,树枝生物量占总量百分比先减小再增大,分别为7.46%、6.73%和6.80%,与树干生物量变化趋势恰恰相反。
树叶生物量所占比例为3.95%~4.27%,各间伐强度均使树叶生物量占比减小,并且随间伐强度增加,变化趋势与树枝基本相同,这是因为树叶具有独特的光合作用功能,当树叶量多时,光合作用固定碳也多,支持枝的生长,枝生长延伸的同时也为叶提供更多的生存空间,使树叶量增多。本研究树叶生物量随间伐强度增大呈“V”型变化趋势。T15样地生物量最小,为1.17 t·hm-2,与CK样地相比减少了7.14%。T35样地树叶生物量最大,为1.54 t·hm-2,比CK样地增加了22.22%。
树皮生物量所占比例为2.40%~2.58%,间伐处理对树皮生物量无显著影响,且间伐对树皮生物量占总量百分比影响较小。
3.3 不同抚育间伐强度下杉木各器官含碳率
由表4可知,在杉木各器官中,树干含碳率几乎均高于其他组分,并且树干占比最大,因此树干是碳的主要载体。云杉、冷杉树干含碳率均在T25样地最大,分别为52.27%、49.50%;含碳率受到树种本身遗传特性的影响以及各器官功能影响,因此在同一地区相同间伐强度处理下,不同树种各器官含碳率亦有所差异,并且杉木的枝、叶、皮含碳率随着间伐强度的增大无明显规律变化。
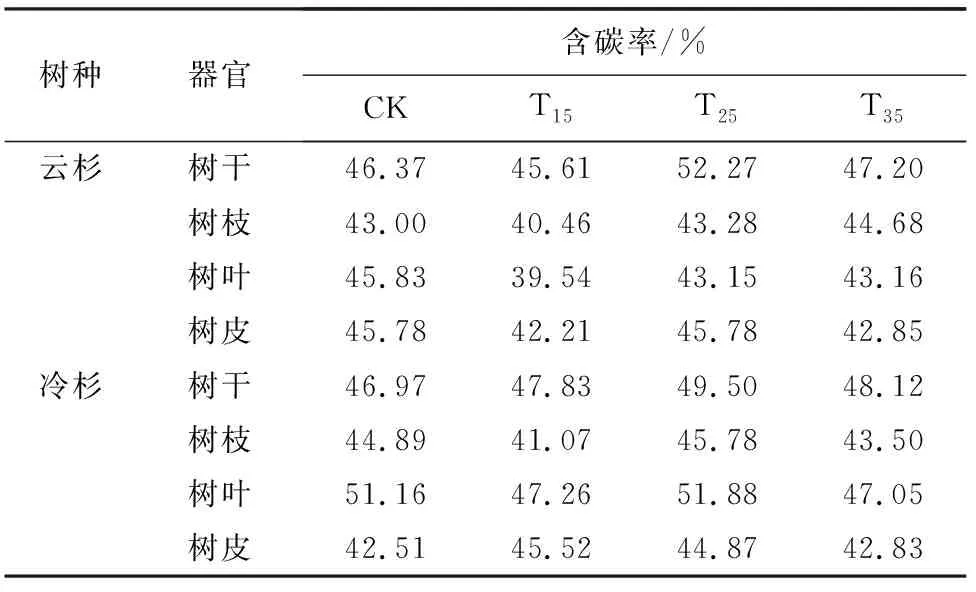
表4 不同间伐强度下林木各器官含碳率
3.4 抚育间伐下林木各器官碳密度
由表5可以看出,在T15、T25、T35样地,地上部分碳密度随着间伐强度的增大先增加再减小,其中T25样地最大,为CK样地的1.42倍,T35样地其次,为CK样地的1.28倍,而T15样地最小,与CK样地相比减少了8.06%,其中CK、T15样地间碳密度无显著差异,T25、T35样地间碳密度无显著差异。不同间伐强度下杉木碳密度在乔木各器官的分配特征一致,由大到小均为:树干、树枝、树叶、树皮。
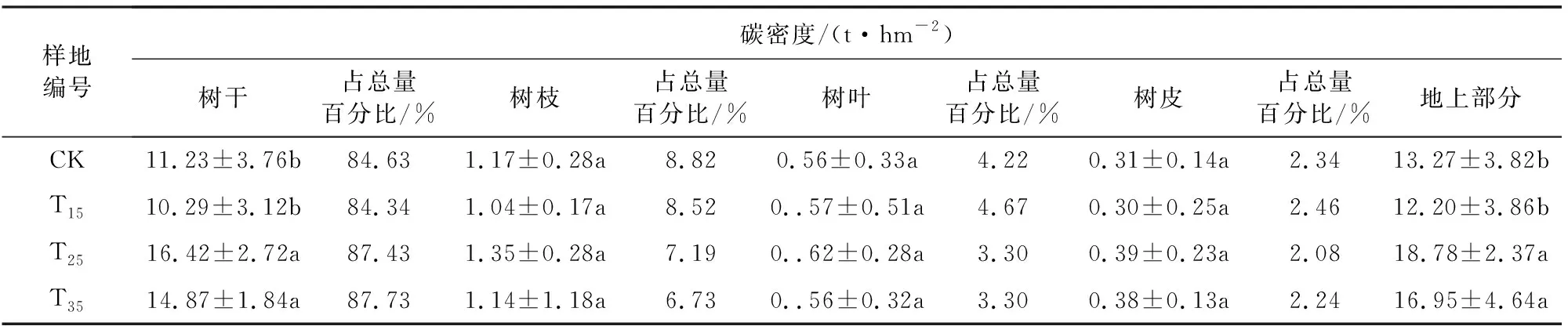
表5 不同间伐强度下杉木碳密度(平均值±标准差)
树干是碳的主要载体,碳密度约占总量的84.34%~87.73%。随着间伐强度增加,树干碳密度所占比例先减小再增大。这是由于间伐强度增加,保留木间竞争减少,稀疏后的林分为保留木提供了较大的上层空间,有利于树枝和树叶的延伸,因此树干碳密度占总量的比例减小,树枝和树叶增大,而间伐强度过大增加了林冠的暴露面积,使林内小气候变化显著,不利于林冠生长,因此树叶树枝所占比例又呈减小的趋势。T15样地的树干碳密度比对照样地减少了8.37%,而T25、T35样地树干碳密度为对照样地的1.46、1.32倍。
树枝碳密度在T25样地最大但与其余样地间无显著差异;树叶碳密度在各样地间无显著差异。树枝碳密度约占总碳密度的6.73%~8.82%,树叶碳密度约占总碳密度的3.30%~4.67%,虽然树叶和树枝碳密度远小于树干,但其具有重要功能,是碳的储存不可缺少的器官,应适当培育,将其调整至最佳状态,有利于森林碳汇。
树皮碳密度占总碳密度的2.08%~2.46%,但各样地间差异不显著。树皮碳密度虽少,但其具有重要功能,可防寒、防暑、防止病虫害,在植物的韧皮部里排列着一条的管道,树叶通过光合作用制造养料,再通过它运送到根部和其他器官中去。
3.5 抚育间伐下Hegyi竞争指数与平均最近距离
抚育间伐通过改变树种分布位置从而改变单木间的竞争关系。竞争指数表现了目标树胸径间的竞争强度,平均最近距离是指对象树周边最相近4株树距离对象树距离的平均值。运用Hegyi竞争指数和平均最近距离共同反映了样地内的激烈竞争程度。
由表6可知,同CK样地相比,T15样地杉木的Hegyi竞争指数减少了2.05%,而T25、T35样地分别增加了36.07%和14.66%。间伐后T15样地杉木的平均最近距离为CK样地的1.1倍,表明此时的竞争压力较大,而T25、T35样地的杉木的平均最近距离(3.2、3.7 m)均大于CK,表明T25样地林木竞争压力较大。
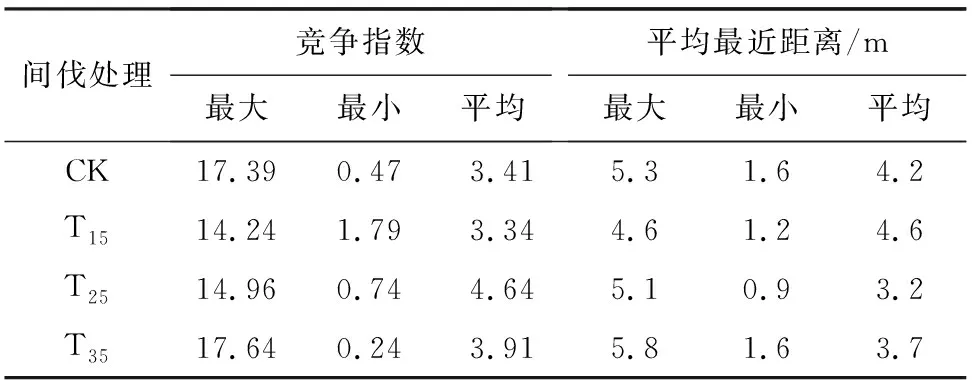
表6 不同间伐强度下林分竞争强度
3.6 竞争与间伐强度对林木各器官碳密度分配特征的环境解释
为探究竞争对小兴安岭东方红林场针阔混交天然次生林树木生长的影响,采用各样地林木Hegyi竞争指数、间伐强度作为环境因子,采用RDA对林木各器官碳密度、胸径、树高和郁闭度等9个指标进行分析,并进行方差分解(图2)。结果表明,Hegyi竞争指数、间伐强度共同解释了碳密度变化的61.3%。竞争指数与第1主轴呈显著正相关(相关系数为0.693 3),竞争指数与H/D和郁闭度的关系呈正相关,相反,竞争指数与地上部分碳密度、树干碳密度、树枝碳密度、树叶碳密度、树皮碳密度、胸径、树高因子呈极显著负相关,说明较大的竞争指数不利于林木碳密度的增长。间伐强度与第2主轴呈显著正相关(相关系数为0.473 2),间伐强度与树叶、树皮碳密度及胸径呈显著负相关,表明间伐强度的增加不利于其增长。
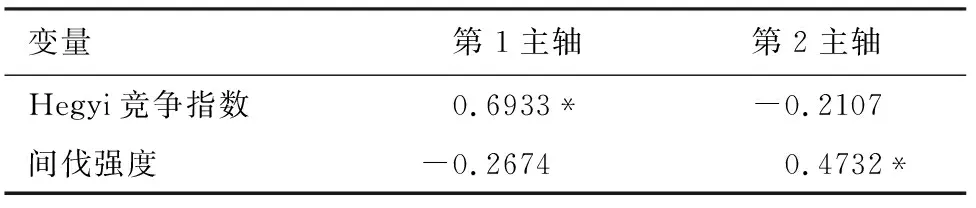
表7 竞争指数及间伐强度与2轴的相关性
4 结论与讨论
小兴安岭带岭林业实验局的天然次生林中杉木经35%强度抚育间伐后林分径阶分布频率偏离正态性,且15%和25%抚育间伐强度的小径木比例增加,林分更新效果好,而35%间伐强度释放竞争压力,使中大径级林木比例增加;间伐改变了林木各器官生物量的分配,使树干生物量占比整体增大,且在间伐强度为15%、25%和35%时树干生物量所占比例呈先增大再减小的倒“V”型趋势,而树枝生物量分配规律则恰恰相反;试验区以15%、25%、35%强度抚育间伐后,与对照样地相比林分生物量均增加,但间伐强度为15%时间杉木碳密度小于CK,间伐强度为25%和35%时杉木碳密度大于CK。间伐强度为25%时杉木恢复较好,适当的竞争压力有利于林木加速生长,碳密度与对照相比显著增加。所以在间伐8 a时,25%间伐强度有利于小兴安岭天然次生林中杉木碳储量的恢复。
不同研究区域生长着不同的树种,以不同强度进行抚育间伐,对林分的群落分布造成的影响也不尽相同。间伐会改变林木的径阶分布[2,17],本研究中对照、T15、T25样地的杉木径阶频率分布呈正态性,而T35样地中杉木径阶分布曲线偏离正态性。T35样地优势树种的平均胸径明显高于其余间伐样地,可以推断该样地抚育间伐8 a后释放保留木间竞争压力,促进了天然次生林林分生长。
间伐强度的大小直接影响被移除林木个体数量,从而影响林分密度的大小,最终导致林分生物量及碳密度的变化。间伐降低林分密度,但是却能增加单株生物量,所以不一定就会减少林分生物量,这与间伐之后的生长年限有关。林分密度大的样地,林木个体生长受有限冠幅的限制,使得供给林木生长的光合同化物减少,林分生物量较小。林分密度较小的样地,林木个体生长显著增加,林分生物量随之增大。小兴安岭针带岭林业实验局的天然次生林中杉木间伐8 a后,间伐强度为15%、25%和35%样地地上生物量大于CK,说明该间伐强度会一定程度上增加林分生物量,这与成向荣等[16]、J.H.Gillisetal[17]研究成果一致。间伐虽移除部分杉木使杉木生物量降低,经过8 a时间恢复,25%抚育间伐强度林分密度适中,林木个体生长显著增加,生物量及碳密度均最大。而间伐强度较低的样地林分密度大,林木个体生长受有限冠幅的限制,使得供给林木生长的光合同化物减少[18],林木个体生长缓慢,最终会导致生物量增加缓慢。碳密度在林木各器官的分布排列一致,由大到小均为:树干、树枝、树叶、树皮。杉木树干的碳密度在间伐强度为15%、25%和35%时,随着间伐强度的增大而先增加再减少,合理的间伐强度有利于杉木树干碳储量的累积,其主要原因是间伐改善林木生长空间,促进了保留木树高和胸径的快速生长。
竞争是植物群落中普遍存在的现象。在生长发育的过程中,植物会与附近的个体争夺水资源、光资源及养分资源等[19]。本研究中T25样地生长情况较好,存在较多的中径级杉木,因此同CK相比,林木的Hegyi竞争指数增长较大。T15样地杉木平均最近距离小于CK,而T25、T35样地的杉木的平均最近距离均大于CK,说明随着间伐强度的增大林木竞争压力增大。RDA结果表明,竞争指数与第1主轴呈显著相关,随着竞争压力增大,胸径和树高逐渐减小,而树高胸径比则反映了植株径向生长策略,当林木承受竞争压力较大时,其径向发展会受到严重的影响和限制,郁闭度也随着竞争指数的增大而增大,这些与段梦成等[3]研究结果一致。地上部分碳密度、树干碳密度、树枝碳密度、树叶碳密度、树皮碳密度、胸径、树高聚集在第1轴反方向上,说明较大的竞争指数不利于其增长,这与周文嵩[20]的研究结果相似。间伐强度与第2主轴呈显著正相关性,间伐强度的增加不利于与树叶、树皮碳密度的累积及胸径增长。竞争因子与间伐强度因子均会不同程度影响碳密度变化。而在分析结果中有未被解释的部分,这表明其他因素也会影响森林碳密度,如气候、光照和土壤因素等[21]。
猜你喜欢
绿色天府(2022年9期)2022-11-19
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
绿色科技(2022年8期)2022-05-25
现代园艺(2021年23期)2021-12-01
桉树科技(2021年3期)2021-09-02
西北园艺(果树)(2021年3期)2021-06-16
绿色科技(2021年24期)2021-02-21
绿色科技(2020年15期)2020-10-14
农民致富之友(2020年8期)2020-05-11
