
贵州普安四球茶(Camellia tetracocca Zhang)种质资源的遗传多样性分析
2022-02-04 11:51陈艳艳尹世华王莉飞黄晓霞程小毛
江西农业大学学报 2022年1期
罗 雯,陈艳艳,尹世华,王莉飞,黄晓霞,耿 芳,程小毛
(西南林业大学 园林园艺学院/云南省古茶树资源保护与利用研究中心,云南 昆明 650224)
【研究意义】近年来,随着茶叶消费观念由求量向求质转变,古茶树逐渐受到重视,政府及茶农开始对优质的古茶树种质资源进行保护性开发和种植推广[1]。四球茶(Camellia tetracoccaZhang)属于山茶属茶亚属Subgen.Thea 茶组Sect.Thea 五室茶系Ser.Quiquelocularis 中的一种,又名炒青茶,是1981 年张宏达先生发现的新种,集中分布在贵州省普安县海拔1 700~1 950 m 的森林中,为贵州特有种[2]。2009 年袁茂琴等[3]通过野外调查比较了大厂茶和四球茶的花、叶、果及地域范围的差异,提供了四球茶作为一个新种从大厂茶里分离出来的形态依据。同时四球茶具有高抗寒性,能够忍受-6.0 ℃以下的低温,可作为远缘杂交、基因转移聚合等茶树育种材料,是实现突破性育种和选育优良茶树品种不可替代的基因源[4],故四球茶在遗传学上具有较高的保护和研究价值。【前人研究进展】目前,对四球茶的研究多见于对其生理[4]、生化[5]、种子繁育[6]和土壤[7]等方面。此外,一些学者也陆续开展了四球茶SSR[8]及SNP[9]分子标记的挖掘,而针对四球茶遗传多样性的研究还较少,这对四球茶的种质保存和遗传育种是非常不利的。而四球茶作为具有地方特色的茶种,其丰富的资源在品种发掘、利用和保护上具有非常重要的价值。因此研究四球茶遗传多样性是今后开展育种工作的基础。
【本研究切入点】EST-SSR 标记又叫基于表达序列标签微卫星(expressed sequence tag based simple sequence repeat),除了具有SSR 的特点外,还有引物廉价、通用性好、条带清晰、容易统计等优点,被广泛应用于植物遗传多样性等方面研究[10]。目前EST-SSR 在茶树上的应用主要在亲缘关系及遗传多样性分析方面[11-12]。【拟解决的问题】研究通过EST-SSR 标记方法对普安四球茶进行遗传多样性和居群结构分析,旨在从分子水平上分析四球茶的现状,为四球茶育种亲本的选择、生态适应性等研究提供理论依据。
1 材料与方法
1.1 材料
选取贵州省普安县青山镇普白林场和马家坪141份四球茶树资源作为实验材料,其中野生型古茶树62 株,千年野生型古茶树5 株,栽培型古茶树17 株,栽培型茶树57 株。采集当年生完全展开的幼嫩叶片,放入装有有色硅胶的自封袋中,做好相应标记,带回实验室自然保存以备用。
根据普安县青山镇茶树分布特点,将所采集材料分为野生型古茶树、千年野生型古茶树、栽培型古茶树和栽培型茶树4个居群,其中野生型古茶树、千年野生型古茶树和栽培型古茶树居群的树龄均是经专家调查鉴定。
1.2 DNA提取
采用CTAB 法[13]提取茶树叶片基因组DNA,再经10 g/L 琼脂糖凝胶电泳检测质量和微量核酸蛋白检测仪测定浓度和纯度后,稀释DNA溶液至50 ng/µL左右。
1.3 EST-SSR引物选择
在程小毛等[14]设计的、由生工生物工程(上海)有限公司合成的200 对茶树EST-SSR 引物中筛选出22对扩增条带清晰、多态性高的引物,有关引物的具体信息见表1。

表1 22对EST-SSR引物合成信息Tab.1 22 pairs of EST-SSR primers correlation properties
1.4 PCR扩增及产物检测
参照唐探等[15]的反应体系及反应程序,扩增产物用质量分数为6%尿素变性聚丙烯酰胺凝胶电泳检测,并参照程小毛等[14]的方法进行银染显色,最后放在灯箱上拍照记录。
1.5 数据处理与分析
采用人工读带的方法,将电泳图上清晰的条带记为“1”,同一位置无带或不易分辨的弱带记为“0”,建立原始数据矩阵,利用Data Trans 1.0 软件[16]将0/1 转换为基因型数据类型。利用POPGENE v1.32[17]软件分析其遗传参数。根据遗传相似度值,利用NTsys 2.1[18]软件进行UPGMA 聚类分析。采用GenALEx[19]软件进行AMOVA 分子差异分析及PCA 分析。利用软件Structure 2.3.4[20]进行居群遗传结构分析,设定K值为1~10,MCMC(markov chain monte carlo)为10 000 次,根据似然值最大的原则,在运算结果中选取一个合适的K值。
2 结果分析
2.1 遗传多样性分析
利用22 对EST-SSR 引物对141 份材料进行PCR 扩增,共扩增出72 条多态性带,每个位点扩增出的等位基因数在2~5 个,平均扩增数为3.272 7 个,扩增位点多态率100%;从表中可知,观测等位基因(Na)的数值最小为3.000 0,分别是CsEMS147、CsEMS148、CsEMS152 和CsEMS194,最大 为6.000 0,是CsEMS27 和CsEMS117,平均4.227 3,有效等位基因数(Ne)最小值为1.647 3,是CsEMS152,最大值为3.798 1,是CsEMS117,平均2.787 4。22 对EST-SSR 引物中,Shannon 信息多样性指数(I)在0.684 6(CsEMS152)~1.505 9(CsEMS117),变幅0.821 3,变化范围较大,平均Shannon指数为1.145 4,说明普安四球茶古茶树具有丰富的遗传多样性。观测纯合度(Obs-Ho)范围为0.049 6(CsEMS38)~0.822 7(CsEMS148),平均0.458 7,高于平均期望纯合度(Exp-Ho为0.375 1)。观测杂合度(Obs-He)为0.177 3(CSEMS148)~0.950 4(CSEMS38),均值为0.541 3,期望杂合度(Exp-He)均值为0.624 9,说明扩增位点的平均纯合度略高于平均杂合程度。Nei 基因多样性指数(H)平均值是0.622 7,范围在0.392 9(CsEMS152)~0.736 7(CsEMS117);引物的多态信息含量(PIC)在13.3%~78.69%,平均为64.38%,表明22对EST-SSR引物的多态率较高。
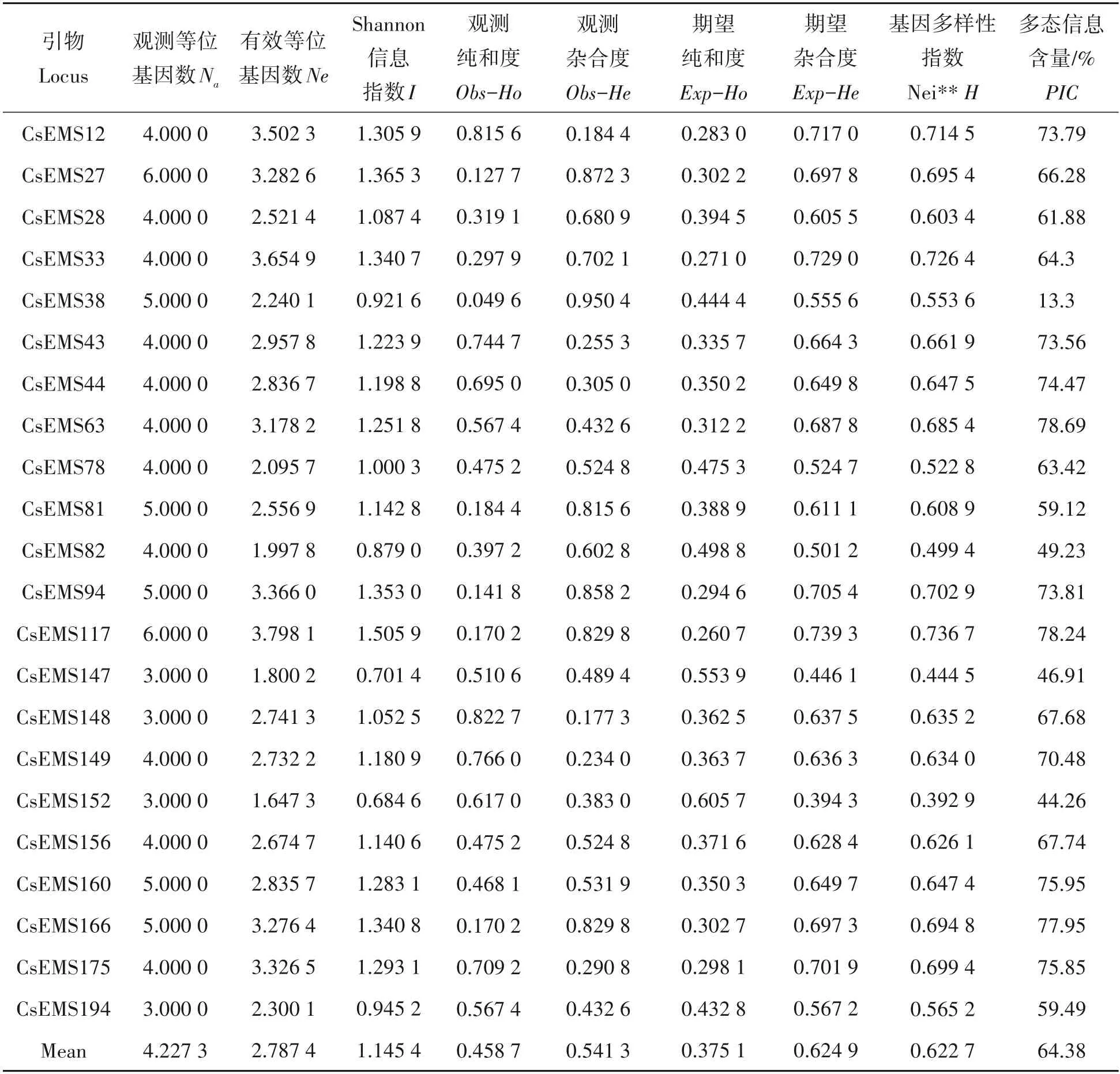
表2 普安四球茶茶树EST-SSR的多样性参数Tab.2 Genetic diversity parameters of Camellia tetracocca Zhang populations in Pu’an based on EST-SSR
不同居群的四球茶遗传多样性的差异见表3。从表中可知,观测等位基因数(Na)分别为4.090 9、3.090 9、3.772 7 和4.090 9,有效等位基因数(Ne)分别为2.551 2、2.641 7、2.747 6 和2.799 6,4 个居群间Shannon 信息指数(I)值大小顺序为:栽培型茶树,栽培型古茶树,野生型古茶树,千年野生型茶树,栽培型的茶树居群的I值为1.133 0,茶树居群遗传多样性丰富,而千年野生型古茶树的茶树居群I值为1.015 5,相比其他类型而言其茶树资源丰富度较小,4 个居群间Nei 基因多样性指数(H)大小顺序为:栽培型茶树,栽培型古茶树,千年野生型古茶树和野生型古茶树。
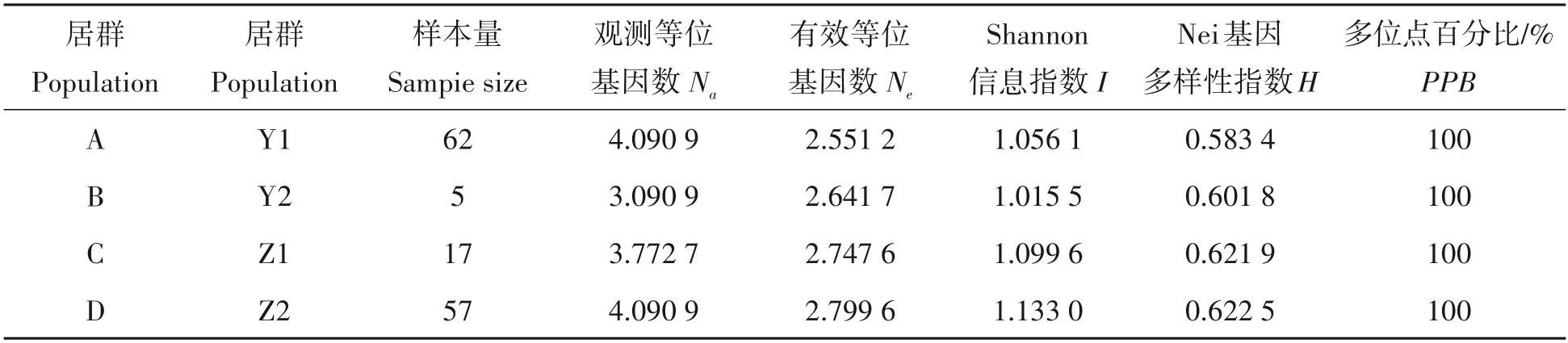
表3 EST-SSR标记在四球茶茶树居群间的遗传多样性Tab.3 Genetic diversity for population of Camellia tetracocca Zhang plants by EST-SSR markers
2.2 遗传分化分析
通过POPGENE 分析得到不同居群的遗传分化结果,本研究居群内的近交系数(Fis)为0.129 3,总居群的近交系数(Fit)为0.170 1,居群分化系数(Fst)为0.046 9,说明四球茶树的遗传变异主要存在于居群内。居群基因流Nm为5.085 3,基因交流丰富。AMOVA 分析表明,四球茶树居群7%的遗传变异发生在居群间,93%的遗传变异发生在居群内,说明普安四球茶的遗传变异发生在居群内(表4)。

表4 普安不同四球茶茶树居群遗传分化参数Tab.4 Genetic differentiation parameters of Camellia tetracocca Zhang populations in Pu’an
普安四球茶居群间的遗传相似度和遗传距离见表5,从表中可以看出各居群间遗传一致度较高,在0.838 0~0.943 8,平均值为0.898 0,说明各居群间相似度较高;相应居群间的遗传距离较小,范围在0.057 8~0.176 7,平均值为0.108 4。其中栽培型古茶树与野生型古茶树居群间遗传相似度最高,达到0.943 8,遗传距离最小,为0.057 8;栽培型茶树与千年野生型古茶树遗传相似度最低,为0.838 0,遗传距离最大,为0.176 7。
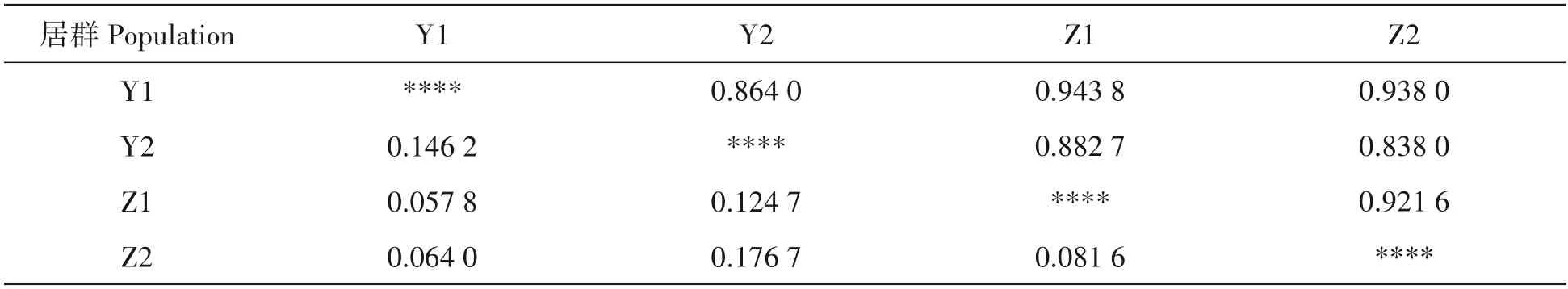
表5 普安四球茶居群间EST-SSR标记的遗传相似性和距离Tab.5 Genetic similarity and genetic distances of Camellia tetracocca Zhang populations in Pu’an by EST-SSR markers
2.3 遗传结构分析
2.3.1 普安四球茶不同茶树居群的聚类分析 通过NTsys 2.1 软件对普安四球茶居群进行聚类分析,当遗传相似度阈值为0.86 时,四球茶居群分为两个亚群,第一个亚群包括居群Y1、Z1、Z2,即野生型古茶树、栽培型古茶树和栽培型茶树居群,第二个亚群为Y2,即野生千年型古茶树居群;其中第一亚群在遗传相似度阈值为0.925时又分为两个亚群,一个亚群是Y1、Z1,即野生型古茶树和栽培型古茶树居群,另一个亚群是Z2,即栽培型茶树居群。从图2中可以看出野生型古茶树和栽培型古茶树遗传相似度最高,栽培型茶树和千年野生型古茶树遗传相似性最小,与POPGENE 分析得到的不同居群遗传相似性和遗传距离结果一致。
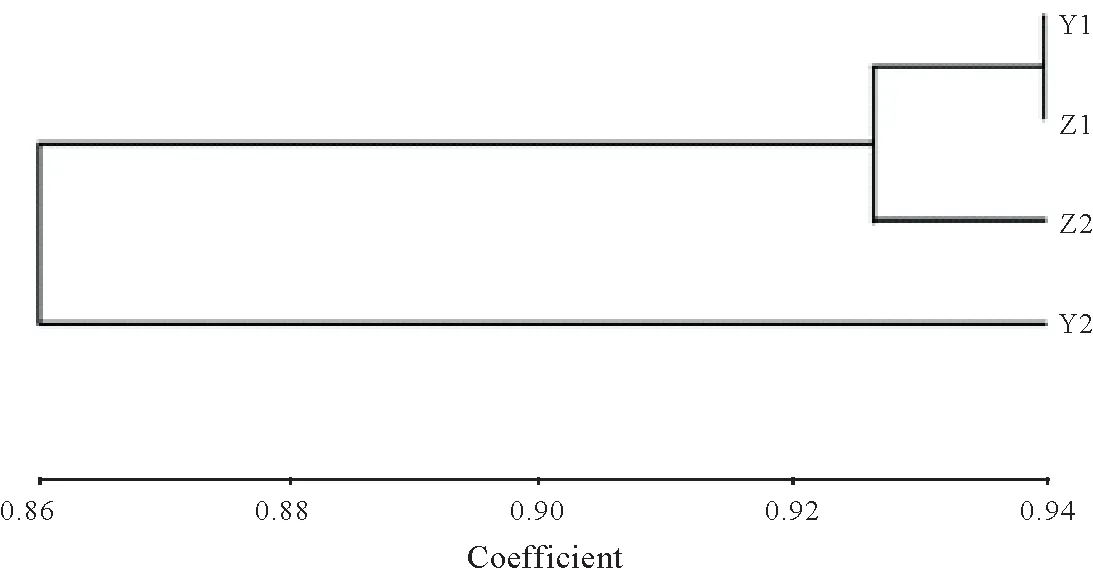
图1 基于Nei遗传相似系数的普安四球茶居群UPGMA聚类分析Fig.1 UPGMA dendrogram for 4 populations of Camellia tetracocca Zhang plants in Pu’an based on Nei’s genetic identity
2.3.2 四球茶树居群结构分析 居群结构分析结果表明(图2),当K=3时,STRUCTURE 将141份普安四球茶种质资源分类为4 个亚群,分类基本与按其生长年限划分的居群相吻合,而且4 个居群间存在频繁的基因交流:紫色柱形是居群A,为野生型古茶树;黄色柱形是居群B,为千年野生型古茶树;绿色柱形是居群C,为栽培型古茶树;红色柱形是居群D,为栽培型茶树。4个居群间存在频繁的基因交流。
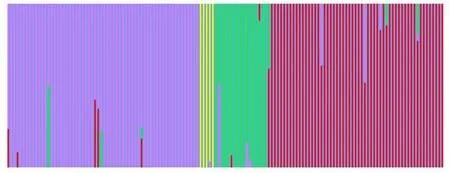
图2 四球茶树4个居群STURCTURE 分析结果Fig.2 Structure analysis with 4 populations of Camellia tetracocca Zhang for the model
2.3.3 基于EST-SSR 的主成分分析 主成分分析(PCA)结果显示(图3),普安四球茶树不同居群间表现出显著差异性,野生型古茶树和栽培型茶树居群较为紧密,栽培型古茶树、千年野生古茶树居群分化明显。其中第一成分占62.8%,第二主成分占31.8%。PCA 分析大部分结果与聚类结果具有相似性,进一步表明所选用的EST-SSR多态性位点具有鉴别普安四球茶树不同居群的能力。

图3 基于EST-SSR标记的主成分分析Fig.3 Principal component analysis(PCA)based on EST-SSR markers
3 结论与讨论
3.1 遗传多样性
本研究中四球茶居群在物种水平上,shannon 信息指数I平均为1.145 4,Nei 遗传多样性指数H平均为0.622 7,多态信息含量PIC平均为0.643 8,物种间遗传多样性丰富,遗传结构差异大。居群水平上,四球茶树居群Na、Ne、I和H的平均值分别为3.761 4、2.685 0、1.076 1和0.607 4,可见四球茶树居群遗传多样性丰富。牛素贞等[9]应用EST-SSR 对贵州具有代表性的茶树品种居群进行遗传多样性研究、和曹烁[10]利用SNP 标记对51 份贵州省野生茶树种质资源的遗传多样性进行分析,结果都表明的贵州茶树种质资源具有丰富多样性,与本研究结果一致。因此,普安四球茶居群遗传多样性丰富的原因可能与茶树是长寿命异交木本植物,而长寿命异交木本植物具有更高的遗传多样性[21];以及贵州具有悠久的植茶历史和多样的生态环境,是茶树起源演化中心之一[22-23]。
本研究中4 个居群间Shannon 信息指数(I)值和Nei 基因多样性指数(H)均表现出栽培四球茶树>野生四球茶树,说明普安四球茶栽培茶树遗传多样性高于野生茶树。这一结果与姜燕华[24]人为选择会导致茶树多态性降低和遗传多样性丧失的观点不一致。但是季鹏章等[25]基于形态研究云南阿萨姆茶、茶、白毛茶和野生茶树大理茶种质资源多样性,结果表明栽培茶树、阿萨姆茶和茶的遗传多样性高于野生茶树大理茶。陈进[26]在对云南不同地区的地方栽培品种与野生近缘种的遗传多样性的研究中,也得到栽培品种遗传多样性并不低于其野生近缘种,均与本研究中普安四球茶栽培茶树遗传多样性高于野生茶树的结果相一致。Zhao等[27]利用14个微卫星标记研究了茶树野生、种植和新近驯化的居群遗传多样性,结果表明野生居群由于过度开发和生境破坏导致遗传多样性丧失。本研究中栽培茶树遗传多样性高于野生茶树,说明普安四球茶遗传多样性受到人为破坏。原因可能是早期当地居民没有意识到茶树的功效和价值,故没有保护野生茶树的意识,甚至为了耕地,对野生茶树肆意损伤、砍伐。另外,由于对纯天然有机茶的需求导致野生茶树价格不断攀升,致使当地居民的过度采摘或随意挖掘部分野生茶树,故造成了一些野生茶树资源的流失。
3.2 遗传分化
Wright[28]指出,Fst值在0~0.05,居群遗传分化较弱;0.05~0.15,居群遗传分化中等;0.15~0.25,表示居群遗传分化较大;当Fst值大于0.25 时,表示分化极大。本研究中,普安县四球茶树居群分化系数为0.046 9,说明四球茶树居群遗传分化较弱,居群间遗传多样性水平低,这与四球茶树居群间的基因流较大有关,在本研究中四球茶树居群间的基因流(Nm)高达5.085 3,足以抵制遗传漂变的作用,防止居群分化。AMOVA 分析表明,普安县四球茶树居群93%的遗传变异来自于居群内,居群间的遗传变异为7%。这与Matsumoto 等[29]关于茶树品种资源的遗传多样性研究结果类似。辛桐[30]对云南山茶(Camellia reticulata)遗传多样性研究得到云南山茶居群内部变异大于居群间变异的主要原因是受到人为的从不同居群进行引种杂交。植物的花粉与种子类型、散布方式和散布范围会对遗传多样性造成影响[31],普安四球茶的散播方式主要是靠种子重力传播,理论上,这样的传播方式因其传播距离有限,其居群间的遗传变异应大于居群内的遗传变异[30],但是本研究的普安四球茶居群内的遗传变异大于居群间的遗传变异,说明在普安四球茶散播过程中,人为因素起了很大作用。
居群的遗传多样性可用平均遗传相似性来度量[32]。本研究中遗传相似度范围为0.838 0~0.943 8,平均值为0.898,大于周萌等[33]基于EST-SSR 标记的云南大茶树遗传多样性分析中遗传相似系数0.41~0.91,平均值为0.65;也大于黄寿辉等[34]对广西部分地区野生茶树遗传关系EST-SSR 标记分析中遗传相似系数在0.53~0.9,平均值为0.71。说明普安县四球茶树居群遗传基础相对比较窄。遗传距离与进化趋异有关,居群或个体间遗传距离越小,表明居群之间的亲缘关系越近;反之,居群或个体间的遗传距离越远,那么,它们之间的亲缘关系越远[35]。本研究中平均遗传距离值为0.108 4,说明普安四球茶居群间亲缘关系近。原因可能是栽培型古茶树是早年间茶农从山里移栽的,来源于野生型四球茶树;栽培型四球茶则是用千年野生型或野生型四球茶的种子进行繁殖的。四球茶野生型古茶树和栽培型古茶树的遗传相似度和遗传距离分别处于最大值(0.943 8)和最小值(0.057 8)。分析原因可能是:茶树属于异花授粉植物,在自然环境中,野生茶树的结实率较低[36]。在古茶树居群中,因自然结实率较低,繁殖后代的能力较弱,在长期的进化过程中,居群内个体的差异性加大,故导致野生型四球茶树居群和栽培型古茶树居群之间的遗传差异加大,遗传分化程度高。
3.3 遗传结构分析
本研究对普安四球茶树不同居群的遗传结构进行了UPGMA 聚类分析、STRUCTURE 分析和PCA 主成分分析:UPGMA 聚类分析表明生长年限对普安四球茶茶树的遗传分化和遗传多样性影响小,造成这一结果的原因可能是不同生长年限的茶树居群由于人为原因不断异交,导致普安四球茶不同居群间茶树基因交流频繁,居群间亲缘关系亦较近[37]。STRUCTURE 软件统计普安四球茶个体聚类虽然多聚于各自居群内,但一些个体还是偏离本居群,造成各柱形交叉明显,这可能是栽培过程中,普安四球茶生境中野生近缘植物的基因渗入栽培类型中,或当地居民直接用当地野生茶培育出新的栽培茶类型,从而导致遗传上的复杂性[38]。PCA 主成分分析表明普安四球茶遗传距离相近,遗传背景复杂,原因可能是长期以来茶树育种多采用从自然授粉居群中单株选择或从优良品种中采集种子进行后代选择的方式[39]。本研究中,栽培型古茶树是当地居民祖辈从深山移植而来,栽培型茶树则是选取生长发育良好的野生古茶树的种子繁殖所得,故形成了栽培茶树复杂的遗传背景。
致谢:云南省万人计划青年拔尖人才项目(80201453)、云南省省级重点学科“园林植物与观赏园艺”建设经费(50097401)同时对本研究给予了资助,谨致谢意!
猜你喜欢
四川蚕业(2022年2期)2022-11-19
农业知识(2022年9期)2022-10-13
茶道(2022年3期)2022-04-27
云南画报(2021年10期)2021-11-24
华人时刊(2021年21期)2021-03-09
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
大众科学(2018年5期)2018-05-21
延河·绿色文学(2016年8期)2016-05-14
