
热带次生林不同林层植物叶片非结构性碳水化合物的季节变化及其对氮磷添加的响应
2022-01-28 07:19陈轶群王艺颖于耀泓温珍杰彭钟通莫其锋
生态学报 2022年1期
陈轶群,王艺颖,于耀泓,温珍杰,彭钟通,周 庆,3,莫其锋,3,4,*
1 华南农业大学林学与风景园林学院, 广州 510642 2 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室, 北京 100091 3 广东鹅凰嶂野外科学观测研究站, 阳江 529631 4 中国科学院小良热带海岸带生态系统定位研究站,茂名 525029
近年来,人类大量施用肥料和使用化石燃料,导致大气氮沉降急剧增加,显著改变了森林生态系统的养分平衡[1]。据相关研究预测,2030年全球大气氮沉降量将达到105.3 Tg N/a[2]。一般而言,沉降的氮可作为养分供给植物的生长需要,但长期、连续、过量的氮输入会降低植物的光合和生长速率[3],引起土壤酸化和生态系统富营养化等一系列环境问题[4]。 我国已经成为全球三大高氮沉降区之一[5],华南地区的氮沉降速率已达到30—50 kg N hm-2a-1,远高于许多欧美国家[6]。由于热带森林发育于高度风化的土壤,土壤中磷有效性较低,持续上升的氮沉降加剧了土壤氮磷比例失衡,极大地改变了森林植物的生长发育及体内碳水化合物的合成与周转过程[7—8]。
植物的生长与发育等是由其体内碳水化合物的动态变化决定的[9]。通常根据碳水化合物在植物体内的存在形式,划分为结构性碳水化合物(Structural Carbohydrates,SCs)和非结构性碳水化合物(Non-Structural Carbohydrates,NSCs)[10]。NSCs主要由可溶性糖和淀粉组成,两者在能量供给上扮演着不同的角色,能够反映植物生理和生长状态及植物对其生存环境的响应情况[11]。研究表明,NSCs含量的变化很大程度上影响着植物的代谢活动及生长过程[12],亦存在明显的季节波动。有研究发现可溶性糖含量一般在休眠季节达到峰值而淀粉含量却在生长季节达到峰值[13]。叶片作为植物对环境变化最敏感的器官,其功能性状能够对环境变化作出快速和准确的响应[14—15]。植物叶片NSCs含量受温度[16]、水分[17—18]及光照[19—20]等环境因子共同影响,呈现复杂的规律性。从热带森林到冷温带森林的研究表明,叶片可溶性糖、淀粉及NSCs含量随着温度和降雨量的增加而降低[16]。植物体内的可溶性糖含量随着土壤水分的增加而降低,淀粉含量却随着水分的增加而升高[17]。也有研究表明,叶片可溶性糖及淀粉的含量随着光强的降低而降低[19]。近年来,国内对森林植物体内NSCs的研究主要集中在不同林分类型[16,18,21]以及乔木物种[22]等方面,通常表现为热带森林植物叶片NSCs含量一般低于温带森林,而乔木一般小于灌木和草本[16]。以往大多数关于NSCs的研究为单次采样或年际间采样,对季节变化研究较少,且没有关注不同林层植物的差异。那么,由于华南地区存在明显的干湿季交替,热带森林中植物叶片非结构性碳水化合物含量的季节变化存在哪些规律?对长期的氮磷养分添加又有哪些响应的差异?不同林层的植物存在哪些响应的差异?鉴于此,本文通过测定华南热带次生林不同林层植物叶片NSCs含量,探究热带森林植物叶片NSCs的季节变化及其对氮磷添加的响应规律,有助于了解热带森林植物对外界环境变化尤其是高氮沉降的响应及适应机制,加深对热带森林生态系统养分限制的认识。
1 材料与方法
1.1 研究区域概况
本研究样地设在中国科学院小良热带海岸生态系统定位研究站内(21°27′49″N,110°54′18″E),地处广东省西南部。该地区主要受热带季风气候影响,年均气温23℃以上,年均降水量约为1700—1900 mm,且干湿季明显,湿季(雨季)集中在4—9月,占全年降雨量的75.8%,干季一般为10月—次年3月。土壤为花岗岩风化而成的砖红壤。该定位站从20世纪60年代起,通过人工引种更新,土地经历了从光裸地-桉树人工林-次生阔叶混交林的演替过程。目前,次生阔叶林林冠层的主要物种有:白车(Syzygiumlevinei)、竹节(Caralliabrachiate)、鸭脚木(Scheffleraoctophylla)等,林下层则以黑嘴蒲桃(Syzygiumbullockii)、九节(Psychotriarubra)、大青(Clerodendrumcyrtophyllum)、紫玉盘(Uvariamicrocarpa)等为主[23—25]。
1.2 试验设计
氮磷添加试验样地始建于2009年10月,在阔叶混交林区域内随机选取植被、地貌、岩性、土壤类型和坡度等基本一致的5个区组,每个区组分成4块10 m×10 m样方,分别为4种不同的处理,包括:1)对照(CK),2)加氮(+N,100 kg N hm-2a-1),3)加磷 (+P,100 kg N hm-2a-1),4)加氮磷(+NP,100 kg N hm-2a-1和100 kg N hm-2a-1)[23]。同一区组不同样方之间设有2 m宽的过渡带且设置隔离板,隔离板埋入地下20 cm。氮磷添加标准主要参考该地区湿氮沉降水平设置。由于热带次生林老成土具有较高的磷吸附能力,所以采用氮磷比1∶1的水平添加。氮处理水平参考本地区湿N沉降水平和鼎湖山氮沉降实验样地进行;由于老成土具有较高的磷吸附能力,所以采用氮磷比1∶1的水平添加磷。施肥处理,每年6次(间隔2个月施用一次)。在每种处理每块样方中,将称量好的试剂溶解在30 L水中后,以背负式电动喷雾器均匀喷洒于林地地表。对照样方也喷洒相同量的水,以避免处理间因外加水不同而造成的影响[23,26—27]。
1.3 叶片采集与测定
试验选择了样本数量充足且分布较均匀的4种植物进行取样和测定,林冠层和林下层植物各2种(表1)。于2019年1月、4月、7月和10月,在不同处理的样地中随机选择健康成熟的3—5个个体进行采样,利用人工采摘与高枝剪结合的方法采集它们的健康成熟叶片20—30片,在封口袋上分别对取样的叶子进行标记。带回实验室后清洗干净,擦去表层水珠后放入烘箱,105℃杀青1 h,然后在65℃烘箱烘干72 h至恒重。之后去除叶柄和主脉,对叶片样品进行研磨封存待用。植物叶片中的可溶性糖用80%的乙醇溶液提取,淀粉用高氯酸法提取,均借鉴Buysse等[28]的测定方法,利用蒽酮-浓硫酸比色法测定可溶性糖和淀粉含量。非结构性碳水化合物(NSC)含量为可溶性糖含量与淀粉含量之和。
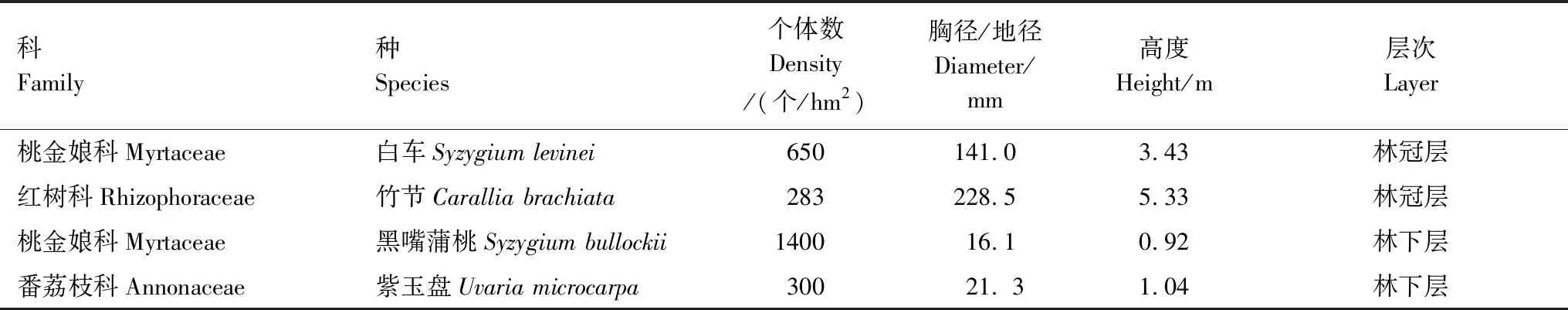
表1 华南热带次生林4种植物的基本特性
1.4 数据处理
所有数据分析均在Excel 2016和IBM SPSS 22.0中进行,利用ANOVA分析比较不同物种、不同处理及不同采样时间植物叶片NSCs含量的差异,对同一林层的植物叶片NSCs含量在干季和湿季间的差异进行配对t检验。图表中数据为平均值±标准误。差异显著水平设定为P<0.05。
2 结果分析
2.1 植物叶片NSCs含量的季节变化
本研究中,植物叶片NSCs含量具有显著的季节差异,且物种与季节的交互作用显著影响叶片可溶性糖和NSCs含量(表2)。多数植物叶片可溶性糖、淀粉及NSCs含量一般表现为干季高于湿季。具体来说,黑嘴蒲桃、紫玉盘和白车的可溶性糖含量均在4月(湿季)最低(图1),紫玉盘、白车和竹节的淀粉含量均在湿季(4月和7月)较低(图2),而竹节的可溶性糖含量及黑嘴蒲桃的淀粉含量在10月(干季)最低(图1和图2)。黑嘴蒲桃、紫玉盘和白车的NSCs含量均表现为4月(湿季)最低(图3)。可溶性糖与淀粉的比值的变化较为复杂,黑嘴蒲桃的可溶性糖与淀粉的比值在1月和4月较低,但均大于1。紫玉盘和竹节在4和10月较低,且紫玉盘在4月的可溶性糖与淀粉的比值均小于1(图4)。

表2 季节(Se)和物种(Sp)对热带次生林植物叶片非结构性碳水化合物影响的双因素方差分析

图1 氮磷添加对不同植物叶片可溶性糖含量的影响Fig.1 Effects of nitrogen and phosphorus addition on leaf soluble sugar concentrations of different plants同一月份中不同小写字母表示处理间差异显著(P<0.05);NP: N、P组合

图2 氮磷添加对不同植物叶片淀粉含量的影响Fig.2 Effects of nitrogen and phosphorus addition on leaf starch concentrations of different plants
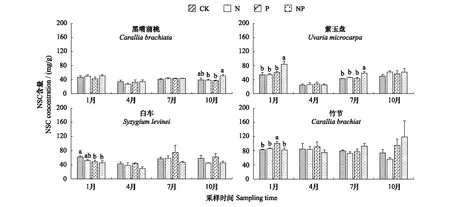
图3 氮磷添加对不同植物叶片NSCs含量的影响Fig.3 Effects of nitrogen and phosphorus addition on leaf Non-Structural Carbohydrates (NSCs) concentrations of different plants

图4 氮磷添加对不同植物叶片可溶性糖含量/淀粉含量的影响Fig.4 Effects of nitrogen and phosphorus addition on leaf soluble sugar/starch ratios of different plants
2.2 不同物种叶片NSCs含量对氮磷添加的响应
植物叶片NSCs含量存在显著的种间差异(表3)。磷添加对叶片可溶性糖、淀粉和NSCs含量具有显著作用,且物种与磷添加对叶片淀粉含量具有显著的交互作用(表3)。黑嘴蒲桃和紫玉盘对N添加的响应较为敏感,白车和竹节则对P添加较为敏感。从整体上看,竹节叶片可溶性糖、淀粉和NSCs含量最高,黑嘴蒲桃最低(图1)。+N和+NP处理均提高了黑嘴蒲桃叶片淀粉含量,且在10月+NP处理显著高于其他处理。除4月和10月+P处理及7月+N处理,N、P添加降低了白车叶片淀粉含量。+P以及+NP处理均增加了竹节叶片淀粉含量(图2)。 N、P添加在7月提高但在4月降低了黑嘴蒲桃的NSCs含量。N、P添加均提高了紫玉盘的NSCs含量。除1月外,+P增加了白车的NSCs含量。1月+P处理的竹节NSCs含量显著高于CK(图3)。本研究中,植物叶片可溶性糖与淀粉的比值存在显著的种间差异(表3)。白车和黑嘴蒲桃的可溶性糖与淀粉的比值高于紫玉盘和竹节,且均大于1。

表3 氮添加(N)、磷添加(P)和物种(S)对热带次生林植物叶片非结构性碳水化合物影响的三因素方差分析
2.3 不同林层植物叶片NSCs对氮磷添加的响应
林冠层植物叶片的可溶性糖、淀粉以及NSCs含量均高于林下层(表4)。氮磷添加增加了林下层植物可溶性糖含量,而减少了林冠层植物可溶性糖含量(表4)。在干季,+N和+NP处理提高了林下层植物叶片淀粉含量,而+P处理降低其淀粉含量,且+NP处理显著高于+P处理;+P和+NP处理增加了林冠层植物淀粉含量,且+P处理显著高于+NP处理,而+N处理减少了其淀粉含量(表4)。在湿季,氮磷添加均增加了林下层和林冠层植物叶片的淀粉含量。在干季,+NP处理下林下层植物的NSCs含量显著高于CK和+P处理。对于林冠层植物,NSCs含量变化情况与干季淀粉含量的变化情况相同。对于林下层植物,氮磷添加会使可溶性糖与淀粉的比值降低,但在干季+P处理会增加其比值。对于林冠层植物,在干季+N处理和+NP处理会增加植物的可溶性糖与淀粉的比值,且+NP处理显著高于CK和+P处理。但在湿季,+P处理显著增加了植物的可溶性糖与淀粉的比值,而+NP处理降低比值且显著低于其他3种处理(表4)。

表4 季节变化对不同林层植物叶片非结构性碳水化合物的影响
另外,不同林层的植物叶片NSCs存在显著的季节性变化,且季节与林层间的交互作用显著影响植物叶片可溶性糖和NSCs含量具有 (表5)。林下层植物叶片NSCs含量和可溶性糖与淀粉的比值均表现为干季高于湿季,林冠层植物淀粉和NSCs含量表现相同,可溶性糖含量和可溶性糖与淀粉的比值表现为湿季大于干季(表4)。

表5 季节 (Se) 和林层 (L) 对热带次生林植物叶片非结构性碳水化合物影响的双因素方差分析
3 讨论
3.1 季节变化对植物叶片NSCs含量的影响
探明植物叶片NSCs的季节变化是掌握树木碳代谢规律的基础。植物体内NSCs在土壤水分变化时的积累或消耗状况取决于物种特定的生长策略[29]。本研究发现季节显著改变热带次生林植物叶片NSCs含量。以往研究表明,经过后期演替存活下来的两种常绿树种体内NSCs的季节性变化较小或没有显著性[30]。成熟树木的NSCs储备对干旱的响应不同,NSCs会促进树木对干旱胁迫的适应[31]。本研究中,植物NSCs在4月处于较低水平,这可能是由于叶片正处于展叶初期需要大量能量投入,所以这一阶段叶片NSCs含量较低。杨芳等[32]在亚热带常绿阔叶林中的研究发现,植物在展叶初期以及中期,可溶性糖、淀粉以及NSCs含量变化缓慢,在当年生叶片的生长进程中,植物叶片可溶性糖含量逐渐增加,而淀粉含量随着叶片的生长成熟逐渐降低。Song等[33]也发现龟甲竹(Phyllostachysheterocycla)叶片NSCs的最低值出现在枝条的快速生长期,而Hoch 等[21]发现叶片NSCs在生长初期含量最高,此后呈下降趋势至10月落叶前最低。可见不同生活型植物对NSCs的利用策略存在显著差异[18]。
另外,本研究发现,热带次生林植物叶片NSCs因林层不同对季节变化的响应也不同。林下层植物叶片NSCs含量和可溶性糖与淀粉的比值均表现为干季高于湿季。研究表明,当植物面对干旱胁迫时,气孔关闭,各项生命活动减弱[34],同时叶片NSCs的转化速率降低,导致生长下降并伴随着叶片淀粉的积累[35]。Würth等[18]对9种植物的研究发现,植物NSCs浓度表现为干季明显高于湿季,并将其归因于干旱限制了植物生长。Liu等[36]研究也表明,干旱导致欧洲山毛榉(Fagussylvatica)幼树叶片NSCs浓度增加,生长速率下降。而本研究发现林冠层植物叶片淀粉和NSCs含量也表现为干季高于湿季,这同样表明干旱胁迫下植物的生长受限,积累了大量的NSCs,低碳输出导致淀粉的暂时积累[35]。而林冠层可溶性糖含量和可溶性糖与淀粉的比值表现为湿季大于干季,这可能是由于当湿季来临时,水分提升可增强植物对养分的吸收和利用,从而提升了光合作用速率[37],合成大量的可溶性糖供给旺盛的生命活动。而林下层植物由于森林郁闭度过高,雨水被林冠层大量截留,导致林下层植物获取的水分较有限,因此雨季带来的水分对林下层植物叶片可溶性糖产生的影响极其有限。不同冠层植物叶片可溶性糖和淀粉的季节动态在一定程度上能够反映出热带森林树木的生长活力和代谢水平[38],这可能是热带森林植物的共存及其对环境的适应而进化出的策略。
3.2 氮磷添加对不同物种叶片NSCs的影响
森林生态系统中,不同生活型植物叶片NSCs含量存在显著的种间差异[18]。刘万德等[39]等研究表明,物种的生物学特性差异是影响NSCs含量变化的主要因素之一。本研究发现,植物叶片NSCs存在显著的种间差异,这与前述研究结果一致。氮素是植物生长和发育的重要营养元素。本研究表明,黑嘴蒲桃和紫玉盘对N添加的响应较为敏感。植物对N添加响应的不同,归因于不同物种对N的利用和分配以及敏感性的差异。研究表明,N添加一般会促进植物生长[40],对植物净光合速率有一定的提升作用,同时可以提高其体内碳水化合物的产量[41],进而影响体内NSCs的积累[42]。但在高N条件下,对N敏感性高的植物光合作用可能被抑制[43]。本研究中N添加一定程度上增加了黑嘴蒲桃和紫玉盘叶片NSCs含量,但降低了白车和竹节的可溶性糖和淀粉含量,说明乔木和灌木对N响应存在显著的差异。霍常富等[44]发现N添加可以增加水曲柳(Fraxinusmandshurica)叶片可溶性糖和淀粉含量,而王雪等[45]报道N添加降低了大针茅(Stipagrandis)和羊草(Leymuschinensis)叶片的可溶性糖含量,其他研究也发现云杉(Piceaasperata)的淀粉含量因氮添加而减少[46]。显然,物种的生物学属性差异是造成它们体内NSCs对氮添加的响应差异的重要原因。
本研究中,白车和竹节叶片可溶性糖、淀粉和NSCs含量对P添加的响应较为敏感。研究表明,P素已成为该热带次生林植物生长限制的关键因子[24,42]。随着P处理年限的增加,植物仍在继续吸收P供给自身生长,以缓解长期以来的P限制[24]。外源P添加显著影响植物叶片的生长[47]和改变叶绿素含量,从而促进或抑制光合速率[48],这些都会进一步影响植物体内NSCs含量的变化。另外,从整体来看,氮磷添加对植物叶片NSCs有不同程度的提升,但N、P共同添加的提升作用最好,这说明N和P的共同施加缓解了土壤的氮磷失衡,满足了植物生长对氮磷需要的协调性,因而促进了植物的碳水化合物的合成,进而使叶片NSCs含量增多,同时可以把多余的N分配给磷酸酶以增强其活性,并缓解土壤P限制,故长势优于单一的N添加和P添加,这与之前对樟树(Cinnamomumcamphora)[49]及大青(Clerodendrumcyrtophyllum)[25]的研究结果一致。
另外,白车和黑嘴蒲桃的可溶性糖与淀粉比值较高且均大于1,表明可溶性糖占NSCs的比例大于淀粉。这与东北地区植物NSCs含量以淀粉为主的研究结论相反[38],与暖温带植物的生长策略相似[50]。这可能是因为树种在温带地区,树木需要合成更多的淀粉以维持在低温环境正常的生理活动。与之相比,本研究区域年均气温较高,植物并不需要储存过多的淀粉来应对低温环境,而是选择转化为可直接利用的可溶性糖,可以使热带植物在生长中更好地调配及利用碳[51]。
3.3 氮磷添加对不同林层植物叶片NSCs的影响
本研究发现,林冠层植物叶片NSCs含量一般高于林下层,这是因为高大乔木处于林冠顶层,直接获取的光能更多,光合作用通常更强[52],合成的碳水化合物也更多。随着林层高度的降低,光照强度随之降低,郁闭度逐渐增加,林下层植物仅能接收到有限的光能,因此降低了林下层植物光合作用的产物,导致林下层植物叶片NSCs含量较低。另外,林下植物为增强自身竞争能力,将更多的碳用以生长,从而获得更多的生存资源和空间[53]。也有研究表明,耐阴树种比阳性树种具有更高NSCs含量[11]。而在本研究中,林下层植物接触到的光较为有限,虽多为耐阴树种,但NSCs含量较低,与前述研究结果并不一致。这可能是由于本研究中的热带次生林的植物群落已相当成熟,物种间耐阴性的差异并不足以导致NSCs含量上的差异。可溶性糖与淀粉的比值反映了植物的碳利用策略,本研究中林下层的可溶性糖与淀粉的比值小于林冠层,这可能是该层物种将更多的碳用于长期能量储存,以适应长期的荫蔽环境,在以往的研究中也有类似结论[39]。
本研究中,不同林层植物叶片NSCs含量对N、P添加的响应不同,林下层植物对N添加响应较为敏感,林冠层植物对P添加较为敏感。不同林层植物叶片的可溶性糖与淀粉的比值对N、P添加的响应也不同。对于林下层植物,氮磷添加降低了可溶性糖与淀粉的比值,但在干季+P处理提高了林下层植物可溶性糖与淀粉的比值,可能是因为该次生林土壤磷有效性较低,补充充足的P素后植物合成大量可溶性糖供给各项生理活动,可溶性糖与淀粉的比值随之升高。对于林冠层植物,在干季N添加会增加植物可溶性糖与淀粉比值,但在湿季P添加显著增加了植物的可溶性糖与淀粉的比值,说明水分与P添加对植物可溶性糖与淀粉的比值有显著的交互作用,水分作为植物生长发育的重要影响因子,其变化必将对植物生理生态过程产生显著影响[54]。虽然N、P添加是影响林层间NSCs差异的因素之一,但光照、水分等环境条件对其影响不可忽视,且林层不同优势种的生理差异也是导致不同林层植物叶片NSCs差异的原因之一。
4 结论
本研究发现,热带森林植物叶片NSCs具有显著的季节动态变化特征,同时也存在显著的种间差异。不同植物对氮磷添加响应的敏感度存在差异,林下层植物黑嘴蒲桃和紫玉盘对N添加的响应较为敏感,而林冠层植物白车和竹节则对P添加较为敏感。林冠层植物叶片的可溶性糖、淀粉以及NSCs含量均高于林下层。另外,不同林层植物对季节变化的响应也不同,林冠层可溶性糖含量为湿季高于干季,林下层植物NSCs含量则表现为干季显著高于湿季。本研究表明,植物所处的生长环境决定其体内碳利用策略,关于不同林层植物非结构性碳水化合物的季节变化及其对氮磷添加的响应,其潜在的机制仍需要进一步深入跟踪研究。
猜你喜欢
今日农业(2022年14期)2022-11-10
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
装备环境工程(2019年9期)2019-10-24
中学生数理化·教与学(2017年4期)2017-04-22
中国医学影像学杂志(2015年9期)2015-12-15
新高考·高一物理(2015年4期)2015-08-20
小小说月刊·下半月(2015年11期)2015-05-14
中国烟草学报(2012年1期)2012-04-09
现代农业科技(2009年19期)2009-03-20
