
高体(Seriola dumerili)线粒体全基因组测定及结构特征分析*
2022-01-19 05:49王开杰徐永江崔爱君柳学周
海洋与湖沼 2022年1期
王开杰 徐永江 崔爱君 柳学周 姜 燕 王 滨
王开杰1, 2, 3徐永江1, 2①崔爱君1, 2柳学周1, 2姜 燕1, 2王 滨1, 2
(1. 中国水产科学研究院黄海水产研究所 山东青岛 266071; 2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 山东青岛 266237; 3. 浙江海洋大学国家海洋设施养殖工程技术研究中心 浙江舟山 316022)


线粒体DNA (Mitochondria DNA)具有结构简单、高拷贝数, 分子量小、偏母性遗传且进化速度快等优点(Curole, 1999)。鱼类线粒体DNA大小一般为15~20 kb, 多为双链闭合的环状结构, 排列紧密, 不同物种间线粒体基因组存在很大差异, 序列中存在串联重复序列以及少数的散在重复序列(周传江等, 2019), 与其他高等动物相似, 鱼类线粒体基因组也是由13个蛋白质编码基因, 2个rRNA, 22个tRNA, 以及非编码区重链复制起始相关的控制区(D-loop)和轻链复制起始区(OL)组成(Satoh, 2016)。随着DNA测序技术的逐渐成熟, 鱼类线粒体基因组已被作为分子标记广泛应用于鱼类种质资源保护, 群体多态性以及系统进化发育等领域(陈四海等, 2011)。黄小林等(2018)探讨了线粒体基因组在6种篮子鱼()系统发育中的适用性; 程佩琳等(2021)通过线粒体全基因组对6种鲟形目鱼类(Acipenseriformes)进行了种群划分; Bernal-Ramírez等(2003)利用线粒体基因组控制区分析了新西兰金赤鲷()群体遗传结构的稳定性。
1 材料与方法
1.1 实验材料及DNA提取
1.2 基因组测序
委托青岛欧易生物科技有限公司进行测序, 流程如下: 将提取的DNA样品参照TruSeqTMNano DNA Sample Prep Kit试剂盒方法构建文库, 采用Covaris M220超声波破碎仪分割成长度为300~500 bp左右的片段并在两端加接头, 进行桥式PCR扩增, 采用Illumina NovaSeq 6000测序技术对样品DNA进行paired-end测序。
1.3 序列组装与分析

2 结果与分析
2.1 线粒体基因组结构特征
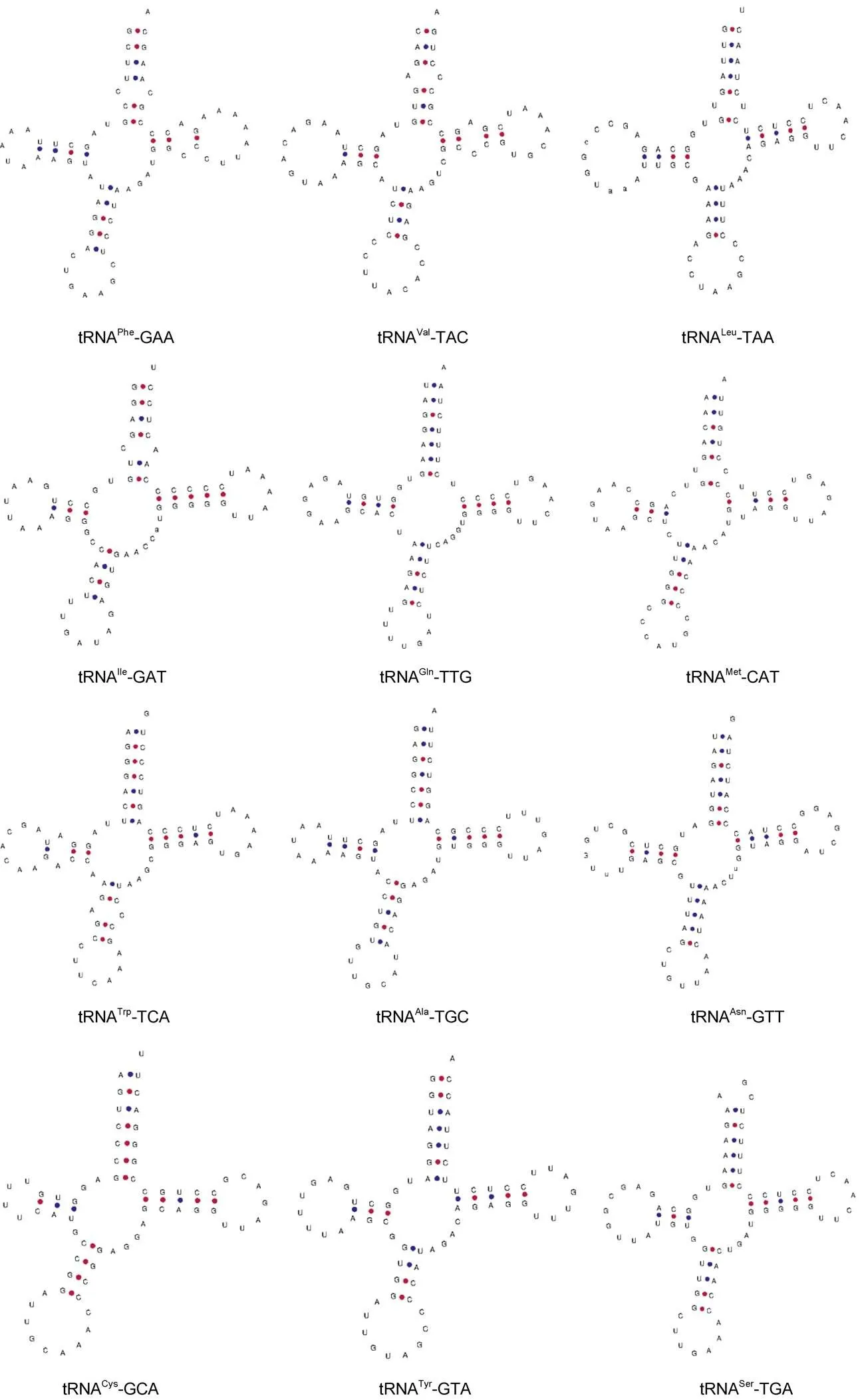
2.2 高体 线粒体基因组rRNA、tRNA及D-loop区结构分析
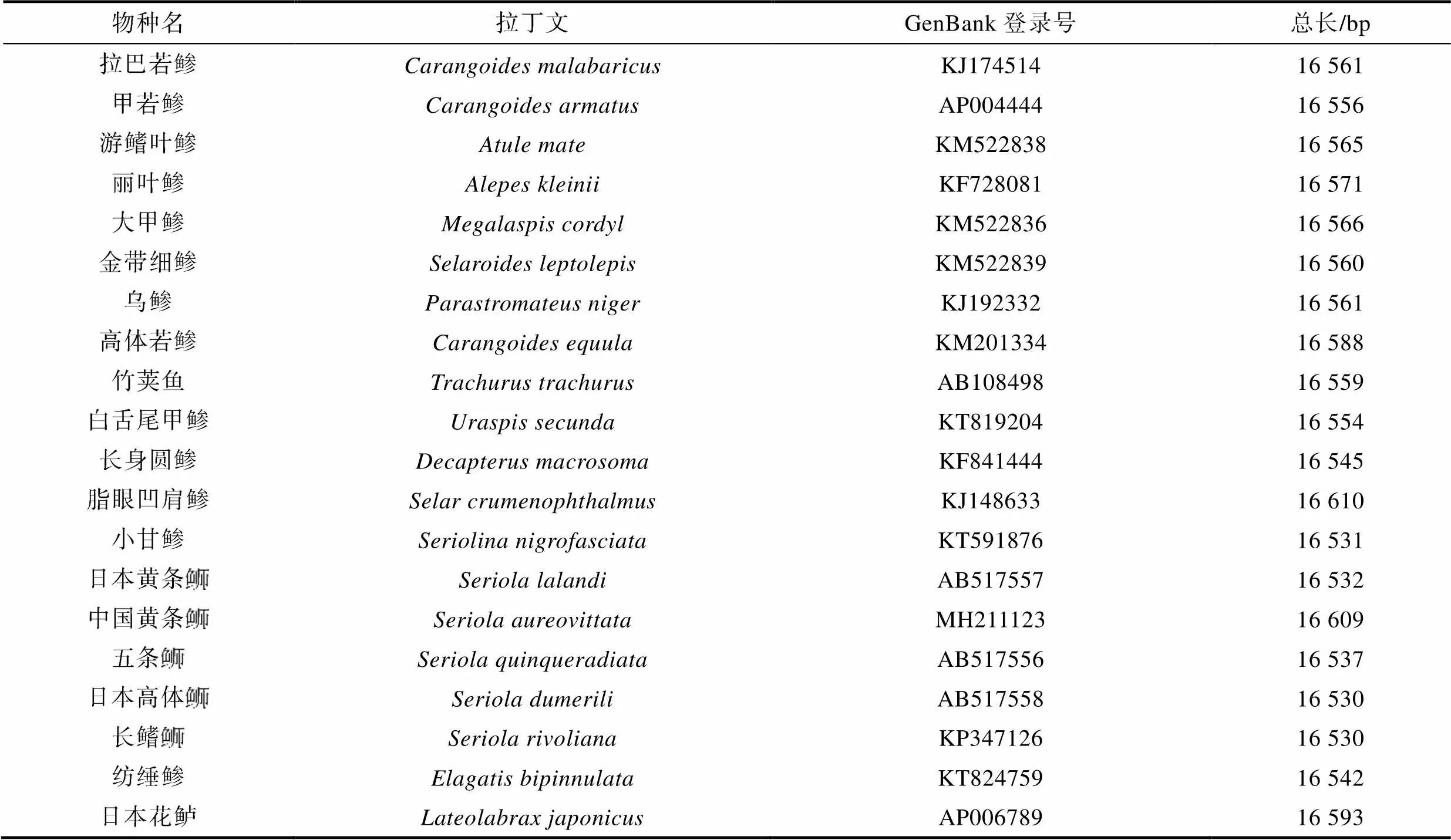
表1 鲹科鱼类线粒体基因组数据来源
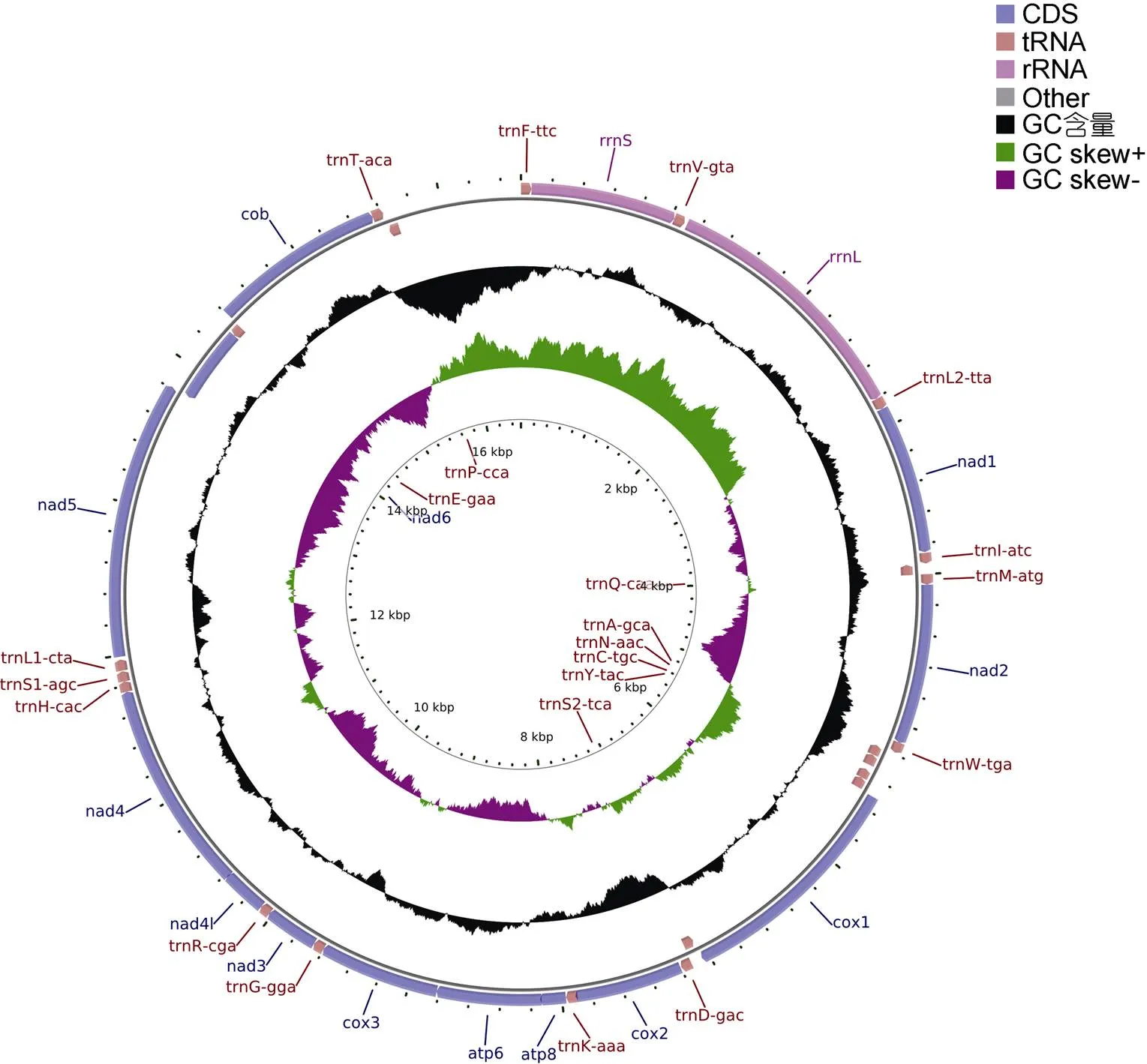
图1 高体 线粒体基因组图谱

表2 高体 线粒体基因组结构特征
22个tRNA的序列总长度为1 552 bp, 长度范围为66~75 bp, 长度最短的为tRNA, 最长的为tRNA和tRNA。其中tRNA和tRNA基因各有两个, 除tRNA、tRNA、tRNA、tRNA、tRNA、tRNA、tRNA、tRNA位于L链外, 其余14个tRNA皆位于 H 链上。22个tRNA结构均可通过tRNAscan-SE在线预测, 除tRNA-GCT基因缺失二氢尿嘧啶臂(DHU臂)外, 其余21个tRNA均含有典型的三叶草二级结构(图2)。
在氨基酸臂中由于C-T转换造成的tRNA、tRNA和tRNA中A-C不配对, 还导致tRNA、tRNA、tRNA和tRNA基因中G-T发生错配。且在氨基酸臂中tRNA-GCT存在1对A-A不配对,tRNA、tRNA、tRNA存在1对C-C不配对,tRNA存在2对T-T不配对。在反密码子茎中也存在由于C-T转换造成的tRNA中A-C不配对。在TΨC茎上, 同样也存在C-T转换现象, 除此之外,tRNA有1对U-U不配对。DHU环也存在G-T错配现象,tRNA有1对A-A不配对, 且tRNA-GCT基因该环缺失。
2.3 高体 蛋白编码基因密码子偏好性分析

表3 高体 13个蛋白编码基因密码子偏好性统计
注: *表示终止密码子; 括号内的字母表示各氨基酸名称的缩写; 表中加粗字体为氨基酸偏好密码子
2.4 中国海域高体 与日本海域高体 蛋白编码基因差异

表4 中国海域高体 与日本海域高体 蛋白编码基因对比
2.5 三种 鱼线粒体基因组比较分析

表5 三种 鱼线粒体基因组结构特点
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋信息技术与应用(2022年1期)2022-06-05
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
生物学通报(2020年11期)2020-10-22
儿童时代·幸福宝宝(2020年9期)2020-09-08
发明与创新·中学生(2019年6期)2019-06-26
中成药(2018年7期)2018-08-04
中国药业(2014年21期)2014-05-26
中国烟草学报(2012年2期)2012-04-09
