
高温胁迫对葡萄叶绿素荧光和光合特性参数的影响
2022-01-18 00:51吴久赟徐桂香李海峰曾晓燕姜建福刘勇翔魏亦农任红松
新疆农业科学 2021年12期
吴久赟,徐桂香,李海峰,曾晓燕,姜建福,刘勇翔,魏亦农,任红松
(1.新疆农业科学院吐鲁番农业科学研究所,新疆吐鲁番 838000;2.石河子大学农学院,新疆石河子 832000;3.中国农业科学院郑州果树研究所,河南郑州 450009)
0 引 言
【研究意义】我国新疆吐鲁番光热资源丰富,是我国重要的葡萄产区,栽培面积达3.68×104hm2,吐鲁番35℃以上的天气年均超过100 d,其中40℃以上的天气年均超过35 d[1-6]。持续高温导致葡萄叶片失水干枯甚至死亡,影响葡萄的光合作用,降低葡萄的产量和品质[7-9]。随着全球平均温度不断升高和持续时间不断延长,高温对葡萄的影响日趋显著[10-12]。研究葡萄对高温胁迫的响应机制,为缓解高温胁迫和探索抗逆栽培途径提供参考。【前人研究进展】光合作用是植物最主要的物质与能量代谢过程,也是植物对高温胁迫最敏感的生理过程[13]。研究发现,高温胁迫会导致Gs和Rubisco酶活性降低,阻抗叶肉细胞的气体扩散,降低植物的光能利用率,并抑制H2O的光解反应和CO2同化能力,影响PSII反应中心活性和电子传递,导致植物不能有效利用吸收的光能,进而使PSII系统结构受损[14-16]。净光合速率(Pn)下降,调运的同化物逐渐增加,最终导致植物体内的能量代谢和物质转化过程失衡,光化学系统PSII被破坏,严重影响逆境诱导基因的表达和调控,造成植株热害甚至萎靡死亡,直接影响葡萄的生长和生产[17-23]。【本研究切入点】美人指葡萄在吐鲁番自然高温条件下仍能保持较高光合效率[6,20],但其具体原因尚不明晰。需要研究高温胁迫对葡萄叶绿素荧光和光合特性参数的影响。【拟解决的关键问题】以美人指葡萄为材料,研究高温胁迫对葡萄叶绿素荧光参数和光合特性的影响规律,为葡萄的逆境栽培和耐热机理研究提供参考。
1 材料与方法
1.1 材 料
试验于石河子大学绿洲生态农业重点实验室进行,供试葡萄品种为一年生美人指(VitisviniferaL.‘Manicure finger’),材料来源于石河子大学综合试验场(85°99′E,44°31′N)。盆栽条件为盆口直径为30 cm、盆高40 cm的白色塑料圆锥形盆钵,盆栽土壤取自试验场葡萄植株根系附近土壤。于2018年9月9日挑选长势基本一致的植株移栽至盆钵中,适应并观察3 d,期间注意保持土壤水分。2018年9月12日将长势一致的盆栽葡萄移入智能人工气候箱(宁波东南仪器有限公司)培养24 h,设置可控条件昼/夜为16 h/8 h,温度为(25±1)℃/(20±1)℃,相对湿度为50%,光照强度为60 000 lx。
1.2 方 法
1.2.1 试验设计
于2018年9月13~16日,挑选长势基本一致的植株放入RDN型人工气候箱,相对湿度为50%,光照强度为60 000 lx。温度设置为35、40、45℃ 3个处理(T),昼/夜时长为16 h/8 h,处理时段为08:00~24:00,其余时间段温度为25℃,以25℃/20℃(昼/夜)为对照(CK)。试验采用单株小区,每个处理3株葡萄,3次生物学重复。
1.2.2 测定指标1.2.2.1 叶绿素荧光诱导动力学曲线(OJIP)及参数
分别于2018年9月13~16日,每日16:00-18:00时,选取第5~7片功能叶,利用快速荧光测定仪Handy-PEA(Hansatech,英国)测定叶绿素荧光诱导动力学曲线(OJIP)及相关参数,每个处理测定3片叶,重复3次。测定前用叶夹暗适应20 min,荧光以3 000 μmol/(m2·s)的饱和光诱导,照射时间为1s。其中OJIP曲线中O点值(F0)表示当所有反应中心完全开放时初始荧光强度,J点值(Fj)表示在OJIP荧光诱导曲线2ms的荧光强度,I点值(Fi)表示在OJIP荧光诱导曲线30ms的荧光强度,P点值(Fp=Fm)表示OJIP荧光诱导曲线的最大荧光强度[21,31]。
1.2.2.2 光合气体交换参数
试验分别于2018年9月13~16日,每日16:00-18:00时,选取第5~7片功能叶,利用LI-6400型便携式光合仪(LI-COR,美国),测定光合速率(Pn),气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等指标,每个处理测定3片叶,重复3次。光量子通量密度为1 800 μmol/(m2·s),参比室CO2浓度为(360±20) μmol/L。
1.3 数据处理
试验数据用平均值±标准偏差(mean±SD)表示,利用DPS 7.05对试验数据进行方差分析,以LSD法进行多重比较并检测差异显著性,以P<0.05为差异显著水平,P<0.01为差异极显著水平;利用SPSS 19.0进行相关分析,用Microsoft Excel 2007做图表。
2 结果与分析
2.1 高温对叶绿素荧光诱导动力学曲线(OJIP)的影响
与对照相比,经过高温胁迫后美人指的OJIP曲线发生了显著的变化,高温胁迫造成叶片最大荧光(Fm)强度下降,但不同温度对OJIP曲线影响不同。其中,不同处理在O-J相的荧光值均高于对照,上升幅度较对照缓慢且无显著性差异,但初始荧光值(F0)均高于对照,说明高温处理可能造成叶片反应中心PSII出现可逆性失活;不同处理在J-I相变化差异显著,其中40和45℃的荧光值上升幅度均低于对照,40和45℃的Fj值和Fi值也均显著低于CK;而35℃J-I相上荧光值升幅度仍高于对照,Fj值和Fi值均高于CK;与对照相比,在I-P相上35℃处理的大部分荧光值都高于CK,其中Fp低于CK,40和45℃处理的荧光值均显著低于CK。高温对OJIP曲线变化趋势影响的规律大体相同,高温导致荧光值降低,且温度越高影响越显著。图1
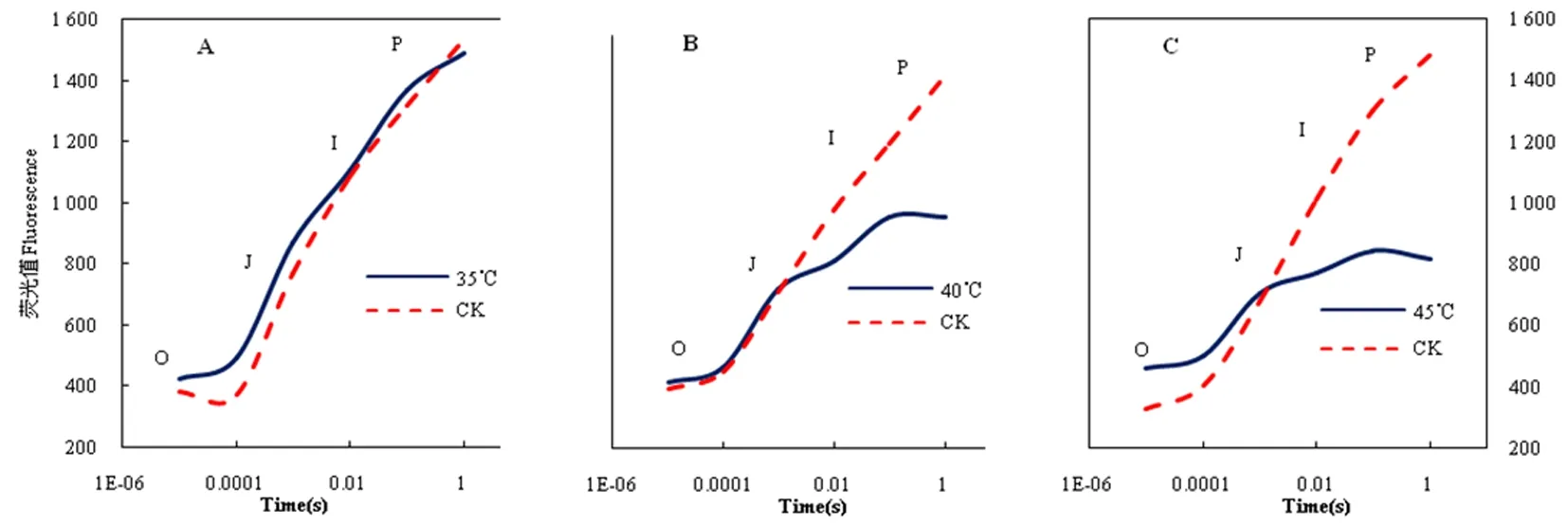
图1 不同温度下OJIP曲线变化Fig.1 The influence of OJIP curve under different temperature conditions
2.2 高温对叶绿素荧光参数的影响
2.2.1F0、Fm、Fv/F0、Fv/Fm
研究表明,45℃处理的F0最高,极显著高于CK,其次是35和40℃,但与CK差异不显著,45℃高温条件下囊体膜受损最严重。45和40℃处理时Fm极显著低于CK和35℃,其中45℃处理的Fm最低,40℃处理次之,CK的Fm最高,反映出温度越高,Fm值越低,热害程度越高。高温条件下Fv/F0随着温度升高而降低,45℃处理的Fv/F0最低,35℃最高,均显著低于CK,且各处理间差异显著,其中45℃和40℃处理的Fv/F0极显著低于CK,说明高温抑制了PSII反应中心的活性,且温度越高抑制越明显。各处理Fv/Fm值为0.48~0.73,均低于CK(0.77),其中45℃和40℃处理Fv/Fm值极显著低于CK,35℃时Fv/Fm值与CK差异不显著。同时,与对照相比各处理Fv/Fm值分别下降了5.19%,27.27%和37.66%,高温对葡萄光能转换效率均有不同程度的抑制作用,温度越高抑制作用越明显。表1

表1 不同温度下葡萄叶片F0、Fm、Fv/F0、Fv/Fm变化Table 1 Effects of high temperature on F0, Fm, Fv/F0, Fv/Fm of grape leaves
2.2.2PIABS、Vj、ABS/RC、DI0/CS0
研究表明,高温处理下PIABS为0.18~1.56,均显著低于CK(5.26),高温影响了葡萄叶片的光能利用效率,导致光合活性降低,且温度越高影响越显著;Vj代表J点的可变荧光强度,反映了QA-~QB之间的电子传递情况和QA-的积累情况,高温处理下Vj为0.52~0.73,均极显著高于CK,高温胁迫造成Vj显著上升,且温度越高上升越明显;ABS/RC表示PSII反应中心吸收的光能,高温胁迫下,各单位反应中心吸收的光能ABS/RC值为1.93~3.10,均高于CK,其中45℃处理与CK差异极显著,反映其温度越高,反应中心吸收的光能ABS/RC值越高;DI0/CS0反映了PSII反应中心以热能消耗能量的比率,与对照相比,高温下各处理DI0/CS0差异不显著,其中45℃处理DI0/CS0最高,为447.31,极显著高于CK,40℃以下热耗散比率与CK差异不明显,45℃处理时,热耗散比率明显升高。表2

表2 不同温度下葡萄叶片PIABS、Vj、ABS/RC、DI0/CS0变化Table 2 Effects of high temperature on PIABS, Vj, ABS/RC, DI0/CS0 of grape
2.2.3TR0/CSm、ET0/CSm、ψo、φE0
研究表明,高温胁迫导致TR0/CSm、ET0/CSm、ψo、φE0均明显下降,且温度越高影响越显著,其中40和45℃处理的TR0/CSm值下降极显著,高温下各处理的ET0/CSm值均显著低于CK,其中40和45℃时ET0/CSm值极显著低于CK,且45℃处理的ψo值仅为0.267,φE0仅为0.13,均极显著低于CK。表3

表3 不同温度下葡萄叶片TR0/CSm、ET0/CSm、ψo、φE0变化Table 3 Effects of high temperature on TR0/CSm, ET0/CSm, ψo, φE0 of grape leaves
2.3 高温对葡萄光合气体交换参数的影响
研究表明,高温导致Pn均有不同程度的下降,其中35和40℃处理与CK无显著差异,但45℃处理下Pn仅为1.66,极显著低于CK,高温胁迫会导致Pn下降,且温度越高Pn下降越显著;高温胁迫下,气孔导度(Gs)均显著低于CK,高温会导致葡萄叶片Gs显著降低,且温度越高越显著;Ci随温度升高而升高,其中Ci为35℃时低于CK,40℃时约等于CK,45℃时则高于CK,且45℃时Ci显著高于35和40℃,高温胁迫下温度越高Ci上升越显著。在高温胁迫下各处理蒸腾速率(Tr)随温度先升高后降低,40℃时Tr显著低于CK,45℃时Tr极显著低于CK。图2
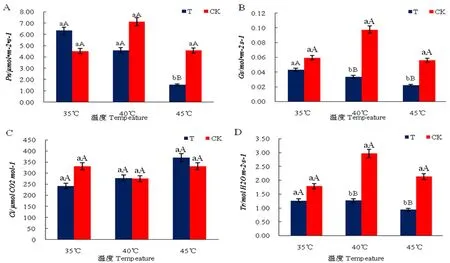
图2 不同温度下葡萄叶片Pn、Gs、Ci、Tr变化Fig.2 Effects of high temperature on Pn, Gs, Ci, Tr of grape leaves
研究表明,Pn与Gs、Ci、Tr均极显著相关(P<0.01),其中Pn与Gs、Tr极显著正相关(0.725**和0.677**),与Ci呈极显著负相关(-0.673**),Gs与Tr极显著正相关(0.969**),Ci与Pn、Gs、Tr均为负相关(-0.673**,-0.128,-0.090)。表4

表4 相关性Table 4 Correlations analysis on Pn,Gs, Ci, Tr of grape leaves
2.4 高温胁迫解除后葡萄光合气体交换参数
研究表明,在高温胁迫16 h,解除胁迫后恢复正常8 h,所有处理的光合作用均可恢复正常。特别是极端高温45℃胁迫解除后恢复正常8 h,美人指葡萄的Pn、Gs、Ci、Tr虽略低于CK,但并无显著差异,在高温胁迫16 h再恢复8 h后,美人指葡萄叶片的光合气体交换参数能够恢复正常水平。表5

表5 恢复8h后葡萄叶片的光合气体交换参数Table 5 Photosynthetic gas exchange parameters in grape leaves after 8 h recovery
3 讨 论
植物对高温胁迫的响应机制受很多因素影响,不同种类植物的热胁迫响应机制也不尽相同[25],轻度高温胁迫通过气孔因子导致植物净光合速率下降,严重的高温胁迫通过非气孔因子导致植物净光合速率下降,高温胁迫下植物净光合速率下降完全是由非气孔因子导致的[30]。研究表明,高温胁迫下葡萄的Pn与Gs、Tr、Ci极显著相关,Gs降低时Ci反而上升,而经过8 h恢复后,美人指葡萄叶片的光合气体交换参数与CK均无显著差异,高温胁迫16 h后导致美人指葡萄Pn降低的原因是非气孔因子导致的[13,15]。
叶绿素荧光诱导动力学曲线能够反映PSII中心原初光化学反应的信息[31],如高温下美人指在O-J相的荧光强度均高于CK,此时叶片接受了大量的光量子[17],而在电子传递过程中荧光曲线(J-I相和I-P相)40℃和45℃的荧光值均显著低于CK,PSII反应中心出现了可逆性失活,与前人研究结果一致[12-13]。研究还发现,35℃时美人指在O-I相的荧光值高于CK,Fv/Fm值与CK无显著差异,较低温度胁迫能短暂提高植物的抗热能力[17,23],但超过40℃时I-P相的荧光强度和Fv/Fm值均显著低于CK(P<0.01),同时PIABS、TR0/CSm、ET0/CSm、ψo、φE0降低,高温会抑制PSII中心捕获光能和电子传递,将电子通过QA-传递到电子受体的比例降低[18],从而导致PSII反应中心的活性和光能转换效率降低[30-32]。
高温导致葡萄Pn、Gs、Tr同时降低,Ci和DI0/CS0同时升高,说明葡萄会通过降低光合作用和提高热耗散来消耗过剩的光能[27-29],也证明Pn的与PSII反应中心活性密切相关[15],而各处理的ABS/RC能量均高于CK,高温下光能主要用于还原QA和热耗散,这可能是葡萄对高温胁迫的一种响应机制[24]。
4 结 论
高温下'美人指'葡萄通过降低PSII的光能吸收、量子产量和电子传递,促进吸收的光能进行热耗散,降低PSII光化学效率,导致CO2同化力降低,进而影响CO2同化和H2O的光解反应。不同温度胁迫对'美人指'葡萄OJIP曲线变化趋势影响的规律大体相同,温度越高导致叶绿素荧光值降低越显著,对葡萄光能转换效率抑制作用也越明显,造成反应中心PSII出现可逆或不可逆失活。可以通过提高PSII反应中心活性,来抵御高温对葡萄光合作用的抑制,如采用遮荫和喷水系统降低环境温度,来缓解高温胁迫对葡萄的伤害,但生产成本较高。
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
中国核电(2021年3期)2021-08-13
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
小资CHIC!ELEGANCE(2018年28期)2018-09-14
女性天地(2016年10期)2017-04-25
分析化学(2016年12期)2017-02-04
小资CHIC!ELEGANCE(2016年15期)2016-07-26
