
郭岩山不同海拔土壤养分对丝栗栲细根碳、氮、磷及其化学计量特征影响
2022-01-14 13:28林君漪柏雨萱林仁忠李新青靳少非郑德祥
江西农业大学学报 2021年6期
林君漪,张 豪,柏雨萱,张 硕,林仁忠,李新青,靳少非,郑德祥*
(1.福建农林大学 林学院,福建 福州 350007;2.福建省顺昌县国有林场,福建 顺昌 353200;3.闽江学院 海洋学院,福建 福州 350108)
【研究意义】生态化学计量学是一门研究生态系统能量平衡与多种化学物质平衡问题的学科[1],通过分析多重化学元素(C、N、P)含量及其计量比值能够进一步探究植被对生态系统结构和功能的影响[2-3]。细根(d≤2 mm)是植物体内运输养分和水分的重要器官[4],它不仅在植物根系系统中最为活跃,且细根快速生长的特性和对养分的高周转速率对于植被维持生态系统中物质循环和能量流动的功能起着重要的作用,对陆地生态系统的生物化学循环具有不可或缺的作用[5-6]。细根也是植物根系中对外界环境变化响应最敏感的部分,具有高度的可塑性。丝栗栲(Castanopsis fargesii)作为我国亚热带地区常绿阔叶林主要建群种,多分布于海拔1 000 m 以下山地常绿阔叶林中。研究丝栗栲细根性状指标可反映该植物生理生态对环境变化响应的关键信息[7]。【前人研究进展】海拔对于植物细根的C、N、P含量及其化学计量比具有影响。李爱琴等[8]对不同海拔杉木成熟林细根功能特征研究表明,随海拔梯度增加,杉木细根C、N 含量表现为先增后减趋势;陈晓萍等[9]对武夷山不同海拔黄山松细根C、N、P化学计量特征研究发现黄山松细根P 含量与海拔梯度存在极显著正相关关系。此外,植物细根的生态化学计量比不仅受海拔梯度的影响,也会受到土壤养分的影响[10]。有研究表明,植物细根P 含量随着海拔梯度的升高与土壤P 含量存在极显著正相关关系[9];但Wurzburger 等[11]研究发现,细根元素指标与土壤C、N、P 含量间不存在显著相关关系。【本研究切入点】海拔梯度对植物细根及土壤养分的影响研究目前主要集中在植物细根功能性状指标、针叶树C、N、P 与其生态化学计量比及土壤养分的变化影响[12-13],而对阔叶树细根C、N、P 含量及其化学计量比以及细根与土壤间C、N、P 含量及化学计量比相关性的研究较少。【拟解决的关键问题】因此本研究将以郭岩山自然保护区的主要树种丝栗栲为研究对象,分析植物细根及土壤C、N、P 元素含量与化学计量特征对海拔变化的响应,揭示丝栗栲天然林细根性状指标的垂直变异规律及其与土壤环境间相互关系,为天然林更新保护提供一定的参考和借鉴。
1 材料与方法
1.1 研究区选择
郭岩山自然保护区是福建省县级自然保护区,坐落于福建省南平市顺昌县,属于南平市与三明市的交接地带,位于27°06′~27°18′N,118°00′~118°30′E,主峰高达1 383.7 m,是顺昌县第一高峰。郭岩山自然保护区地处亚热带地区,属海洋性季风气候,四季分明,年降雨量和日照充足,土壤类型以黄红壤(500~700 m、900~1 000 m)、黄棕壤(700~900 m)为主,郭岩山自然保护区具有典型的福建省中亚热带植被特征,其植被分布具有明显的垂直地带性,区内海拔低于500 m 多以杉木(Cunninghamia lanceolata)人工林为主,海拔1 000 m 以上以肿节少穗竹(Oligostachyum oedogonatum)及小乔木为主,海拔在500~1 000 m 以丝栗栲、木荷(Schima superba)、锥栗(Castanea henryi)和马尾松(Pinus massoniana)为主,丝栗栲为其主要建群树种。
1.2 样地设置
根据研究区内丝栗栲分布状况,分别在600 m、800 m 和1 000 m 共3 海拔梯度下各设置3 个30 m×30 m 的样地。采用相邻网格法将样地平均分为36 个5 m×5 m 的小样方,对每小样方中胸径大于或等于5 cm 的植株进行每木调查样地基本概况详见表。

表1 样地概况一览表Tab.1 Summary of sample plots
1.3 细根及土壤样品采集
在各海拔的每个样地内随机选取5株生长状况良好、树形基本一致的丝栗栲。以丝栗栲树干基部为中心进行细根样品取样,将根系样品置于塑料容器中抖动1 min,从根上分离的土壤团聚体(0.5~5 mm)为根际土,进行自然风干,将收集的细根样品用水轻缓冲洗后过0.8 mm 的网筛,使细根与大部分土壤杂质分离,然后在清水中利用网勺和镊子捡取所有的丝栗栲细根(活根)(直径≤2 mm)。
1.4 细根及土壤样品指标测定
细根及根际土全C、全N 采用碳氮元素分析仪(VarioMaxE,lementalAnalyzer,Germany)测量样品中的单位质量元素含量。细根及根际土全P 的测量步骤为:样品中加入高氯酸消煮后用连续流动分析仪(San++,Skalar,Netherlands)测定。
1.5 数据处理分析方法
采用SPSS 26 软件中的单因素方差分析(one-way ANOVA)分别对不同海拔丝栗栲细根及根际土C、N、P 与生态化学计量比进行方差分析,检验细根C、N、P 含量及化学计量特征在不同海拔梯度之间的差异是否显著;通过Pearson分析不同海拔细根C、N、P含量及化学计量特征与土壤养分之间的相关性。
2 结果与分析
2.1 海拔对丝栗栲细根根际土C、N、P含量及化学计量特征的影响
丝栗栲根际土C、N、P 含量平均值分别为(39.84±3.00)mg/g、(2.23±0.17)mg/g 和(0.53±0.04)mg/g,海拔对根际土C、P 含量存在极显著影响(F=12.780,P=0.000;F=36.476,P=0.000),对根际土N 含量存在显著影响(F=4.204,P=0.021)。丝栗栲根际土C、N 含量均在800 m 海拔处达到最大值,与600 m 海拔之间均存在显著性差异(P<0.05);丝栗栲根际土C∶N、C∶P 平均值分别为(18.52±0.46)、(95.38±7.97),均表现为1 000 m 海拔<600 m 海拔<800 m 海拔,其中C∶N 呈现“倒V 型”变化特征,800 m 海拔与600 m、1 000 m 海拔之间存在显著性差异(P<0.05);丝栗栲根际土N∶P 平均值为(5.34±0.53),随海拔的升高丝栗栲细根根际土N∶P 值显著升高,600 m 海拔与800 m、1 000 m 海拔比值间差异显著(P<0.05),且海拔对丝栗栲细根根际土C、N、P 化学计量比均存在极显著性影响(F=16.409,P=0.000;F=59.757,P=0.000;F=29.780,P=0.000)。由此可见:海拔对丝栗栲细根根际土C、N、P 及其化学元素计量比均存在显著性影响。由于800 m 海拔处林分密度和郁闭度较小,林内光照利用率高,适宜的温度有利于土壤有机质的积累,因而土壤C、N 含量均表现为800 m 海拔处大于其余2 个海拔。其次由于P 元素为典型的沉积性元素,其来源相对固定,大多以成土母质的风化为主,土壤P 含量随海拔梯度升高而降低,可能与该海拔范围土壤的成土母质有关。
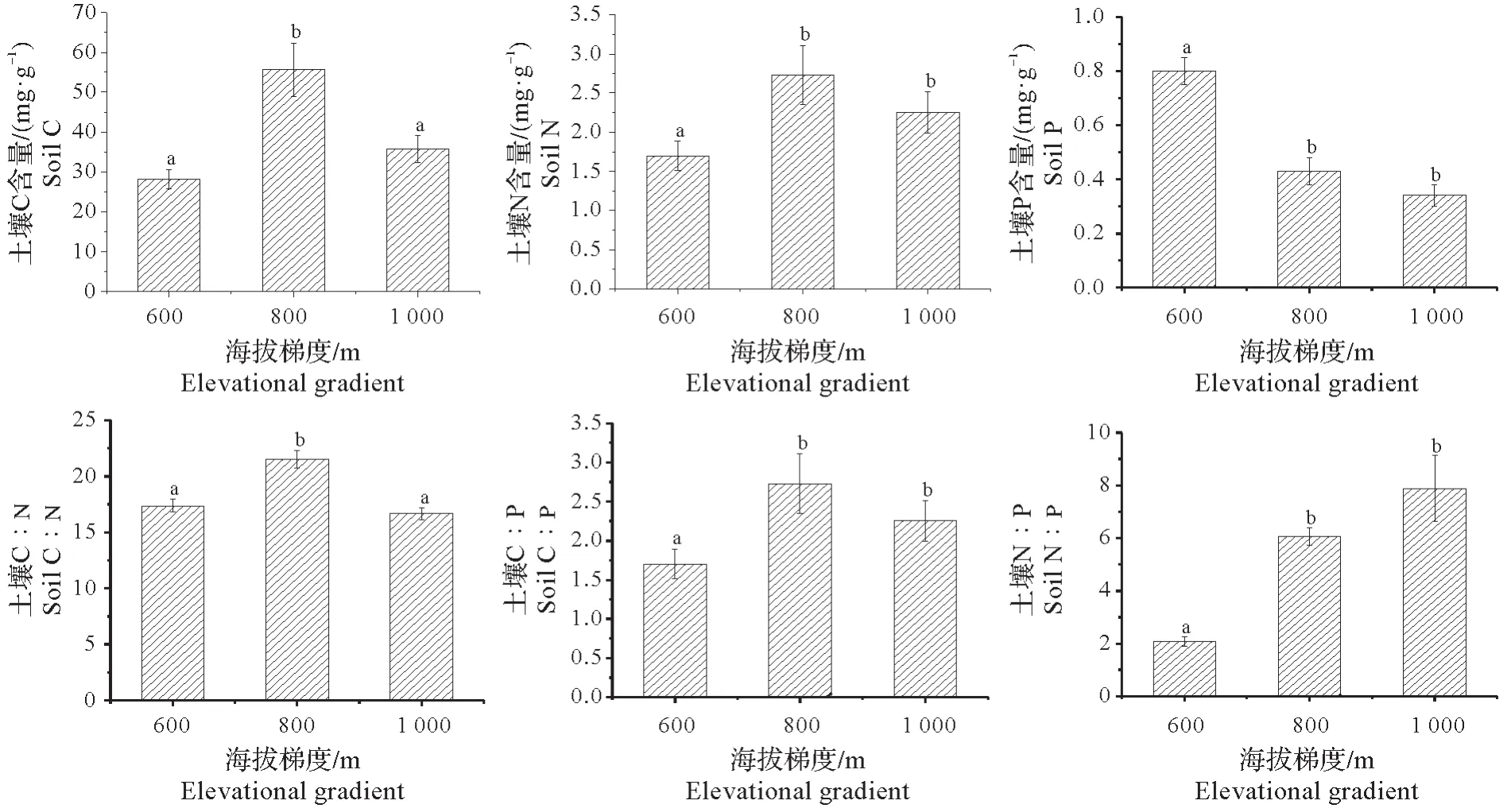
图1 不同海拔梯度下丝栗栲土壤C、N、P含量及其化学计量比变化特征Fig.1 Changes in soil C、N、P content and stoichiometric characteristics in Castanopsis fargesii community under different altitude gradients
2.2 海拔对丝栗栲细根C、N、P含量及化学计量特征的影响
丝栗栲细根C含量平均值为(431.34±3.16)mg/g,海拔对其含量存在极显著影响(F=6.363,P=0.004),丝栗栲细根C含量在800 m海拔处达到最大值,且与600 m、1 000 m海拔之间均存在显著性差异(P<0.05);丝栗栲细根N 含量平均值为(12.81±0.51)mg/g,海拔对丝栗栲细根N 含量有极显著影响(F=10.751,P=0.000),含量变化由小到大依次为海拔1 000 m、600 m、800 m,800 m 海拔与1 000 m 海拔之间存在差异性;丝栗栲细根P 含量平均值为(1.66±0.06)mg/g,海拔对其存在极显著影响(F=17.067,P=0.000),P 含量在600 m 海拔时达到最大且与800 m、1 000 m 海拔P 含量差异显著(P<0.05);丝栗栲细根C∶N平均值为36.77±1.46,各海拔间均无显著差异;C∶P 平均值为(1 122.71±88.36),随海拔的升高丝栗栲细根C∶P 显著增高;丝栗栲细根N∶P 平均值为(8.41±0.47),表现为800 m 海拔数值最高,且与600 m、1 000 m 海拔差异显著(P<0.05)。海拔对丝栗栲细根N∶P、C∶P 均存在极显著影响(F=10.053,P=0.000;F=17.969,P=0.000),对细根C∶N 影响不显著(F=3.198,P=0.050)。结果表明:海拔对丝栗栲细根C、N、P 含量具有较显著的影响,且林分环境对其含量亦有影响,在800 m 海拔地区,由于林分密度小改变了林分内的光照与温度,从而800 m 海拔的C、N 含量高于1 000 m 海拔处,而随着海拔升高,温度降低,林分的生长速率降低,植物对于合成蛋白质的富磷rRNA 的需求下降,从而使800 m、1 000 m 海拔的P 含量低于600 m 海拔。

图2 不同海拔梯度下丝栗栲细根C、N、P含量及其化学计量比变化特征Fig.2 Changes in the C、N、P content and stoichiometric characteristics of fine roots in Castanopsis fargesii community under different altitude gradients
2.3 丝栗栲细根与根际土C、N、P含量及化学计量特征的相关性
对丝栗栲细根与根际土C、N、P及其化学计量比进行Pearson相关性分析,细根C含量与根际土C∶N存在显著正相关(P<0.05);细根N含量与根际土C、N含量之间存在极显著的正相关关系(P<0.01),与根际土N含量的相关系数最大,为0.591;细根P含量与根际土P含量存在0.01水平上的正相关关系(P<0.01),与根际土C∶N、N∶P分别呈极显著负相关(P<0.01)和显著负相关关系(P<0.05);细根C∶N与根际土C、N含量均呈极显著负相关关系(P<0.01);细根C∶P与根际土C含量、C∶N间的关系分别为显著正相关(P<0.05)和极显著正相关(P<0.01);细根N∶P与根际土P含量存在极显著负相关关系(P<0.01),系数为-0.848,与根际土N∶P、C∶P 存在极显著正相关关系(P<0.01),与根际土N∶P 的相关系数最大,为0.613。丝栗栲细根C、N、P 及其化学计量比与根际土其他指标之间未发现显著性相关。可见,丝栗栲细根的化学元素与其根际土化学元素之间存在紧密联系。

表2 丝栗栲细根C、N、P及化学计量特征与细根根际土养分相关系数Tab.2 Correlation coefficient of C,N,P concentrations and stoichiometric characteristics of fine root and rhizosphere soils of Castanopsis fargesii community
3 结论与讨论
3.1 郭岩山丝栗栲细根C、N、P含量及其化学计量比
植物细根组织中的C、N、P 含量对植物生理生态功能具有重要影响,郭岩山丝栗栲细根C 含量平均值为(431.34±3.16)mg/g,与马玉珠等[14]研究的中国植物细根C 含量平均值(473.9 mg/g)相比,其平均值略低,但远高于胡瑞芝等[15]研究的华北落叶松与李爱琴等[8]研究结果中的安徽省天马自然保护区杉木细根C 含量。郭岩山丝栗栲细根N、P 含量平均值分别为(12.81±0.51)mg/g、(1.66±0.06)mg/g,均高于中国植物细根N、P 含量平均值(9.16 mg/g)、(0.954 mg/g)[14],并且超出Yuan 等[16]研究的全球植物细根N、P 含量范围(9.9~11.2 mg/g)、(0.55~0.85 mg/g),此结果与Kerkhoff 等[17]研究发现的多数亚热带和热带地区植物具有高N 含量部分结果一致。丝栗栲细根N 含量与中国及全球植物细根平均值产生差异的原因可能与东南亚热带地区是中国氮沉降较为显著的地区之一有关[18],土壤N 含量及植物吸收N 含量由于大气氮沉降影响显著增加,从而对植物细根N 含量产生影响[19]。本研究中丝栗栲细根P 含量略高于中国及全球植物细根P 含量平均值,与Kerkhoff 等[17]研究发现具有较高N 含量、较低P 含量植物多在热带、亚热带地区出现的结果不一致,其原因可能与采样时间为秋末冬初有关,通常植物在这个季节的生理代谢速率和细根生长速率均较为缓慢,不需要消耗细根中较多的富磷rRNA 来合成蛋白质从而满足自身生长需求。
丝栗栲细根C∶N、N∶P 平均值为(36.77±1.46)、(8.41±0.47),均低于中国植物细根(59.15、14.27)[14]及全球植物细根C∶N、N∶P 平均值(41.41、10.82)[20-21],这是由于丝栗栲细根N、P 含量较高所致;有研究表明N∶P 是指示植被是否受N 或P 限制的重要指标,当N∶P<12 时植被受N 限制,当N∶P 介于12~14 时,植被同时受N 和P 限制或都不受限制,当N∶P>14 时,植被生长受到P 元素限制[22],本研究丝栗栲细根N∶P 平均值为8.41±0.47,说明丝栗栲在一定程度上受到N元素的限制。
3.2 郭岩山丝栗栲细根C、N、P含量及其化学计量比的海拔效应
本研究结果表明,丝栗栲细根C 含量随海拔升高表现出先增后减的变化,且与600 m、1 000 m 海拔存在显著性差异,丝栗栲细根N含量在海拔梯度间存在显著差异,且N含量也呈现“倒V型”变化特征,与李爱琴等[8]和陈晓萍等[9]研究发现细根N 含量随海拔升高而增加的结果不一致。由于800 m 海拔处丝栗栲林分密度和郁闭度较小,缺少树冠遮荫,可大大提高林木光照利用率,温度和土壤环境较优越,有利于地下产物的增加,进而使其800 m 海拔处丝栗栲细根C、N 含量较高,有利于丝栗栲细根的生长。丝栗栲细根P 含量由小到大表现为海拔800 m、1 000 m、600 m,可能是由于600 m、1 000 m 海拔林分密度较大,林内温度较低,林分的生长速率降低,植物对于合成蛋白质的富磷rRNA的需求下降[17],从而使海拔600 m、1 000 m 的P 含量高于海拔800 m。此外,实验结果表明细根P 含量与土壤P 含量呈显著正相关,而土壤P 元素为典型的沉积性元素,其来源相对固定,大多以成土母质的风化为主[10],因而细根P 含量与C、N 含量在垂直带普上的变化特征不一致。这也说明,细根化学元素含量不仅受到海拔、土壤化学元素含量影响,还与丝栗栲所处微环境有关。
海拔对细根C、N、P、C∶P、N∶P变化均具有极显著影响,说明丝栗栲细根化学元素含量在海拔梯度上存在显著变化。细根C∶P、N∶P 在海拔梯度间存在显著性差异,随海拔升高C∶P 表现出先增加后减少的变化特征,这可能与C属于具有相对稳定性的结构性元素[23]且细根P含量受到土壤母质风化程度的影响有关。而N∶P在海拔梯度上表现出先增加后减少的变化特征,这可能是由于在温度较高地区,植物光合速率和吸收速率提高,有利于促进细根对N 元素的吸收[17],而800 m 海拔处丝栗栲林分郁闭度较小,林内温度较600 m、1 000 m 海拔高。根据Garkoti[24]对喜马拉雅山不同海拔的研究,结果显示随着海拔梯度升高,植物根系中N、P的含量降低,而N∶P增加,本研究与上述结果不一致,这表明细根C、N、P化学计量特征除受海拔、土壤养分等因素影响外,还会受树种影响,由于针叶树和阔叶树细根构型不同,导致阔叶树种对土壤养分的吸收速率显著高于针叶树种[25]。Garkoti等[24]选择的研究对象均为针叶树种,而本研究选择的是阔叶树种丝栗栲,因而在不同海拔梯度,不仅需要关注温度、土壤环境的异质性,还需注重物种间的不同特性。
3.3 郭岩山细根C、N、P 含量及化学计量比与细根根际土养分的关联性
土壤是植物生长发育最主要的养分来源,实验对丝栗栲细根C、N、P 含量与土壤养分间相关关系进行研究后发现丝栗栲细根C、N、P含量与土壤养分间存在不同程度的联系。研究结果表明丝栗栲细根C含量与土壤C、N、P含量不具有显著相关性,C元素作为植物的结构性元素,具有相对稳定的特性[23],与陈晓萍等[9]对武夷山黄山松细根研究结果一致。丝栗栲细根N 含量与土壤C、N 元素均存在极显著正相关关系,说明较高C、N的土壤环境更有利于植物细根对于N元素的吸收[17],这与郭炳桥[26]和Burton等[27]的研究结果相似,土壤C、N含量高的地区,细根也具有较高N含量。
丝栗栲细根P 元素含量随土壤P 含量的增加而增加,相关性极显著,而与土壤C∶N、C∶P 呈显著负相关关系,说明相对比于土壤C 元素,郭岩山不同海拔丝栗栲在生长过程中对于N、P 元素的需求可能更大,推测土壤C 含量的提高有利于加强植物细根对于P 元素的吸收[23]。丝栗栲细根C∶N 与土壤C、N 元素含量均存在极显著负相关关系,可能由于大气氮沉降影响,土壤及细根N 含量增加,C∶N相应减少,这与Li 等[28]对关于模拟氮沉降对植物根系性状影响的54 篇论文荟萃研究结果一致。本研究中,丝栗栲细根C∶P 随土壤C 含量增加而显著增加,与土壤C∶N 呈显著正相关关系;细根N∶P 与土壤N∶P、C∶P 具有极显著正相关关系,而与土壤P 含量呈极显著负相关关系,相关系数为-0.848,土壤C∶P 与N∶P 作为土壤养分供应有效性的重要指标[29],有研究表明C∶P 低时,土壤P 有效性较高[1],进而有利于细根对于P 元素的吸收,而土壤P 有效性提高可能造成植物细根N∶P 降低,使植被生长面临N 元素限制。
猜你喜欢
散文诗(2021年22期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
水土保持研究(2020年2期)2020-03-19
环球时报(2019-05-23)2019-05-23
乡村科技(2019年21期)2019-02-21
四川农业大学学报(2018年3期)2018-09-04
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
亚热带资源与环境学报(2015年1期)2015-01-22
郑州大学学报(工学版)(2014年6期)2014-03-01
