
昆虫金属硫蛋白的研究进展
2022-01-11 06:19:10张佳丽周小草魏洪义
生物灾害科学 2021年4期
张佳丽,周小草,罗 妹,魏洪义
昆虫金属硫蛋白的研究进展
张佳丽,周小草,罗 妹,魏洪义*
(江西农业大学 农学院, 江西 南昌 330045)
金属硫蛋白(metallothioneins, MT)具有清除自由基、调节生物体内微量元素浓度、参与重金属的解毒等作用。介绍了MT生物学特性、表达调控生理功能,并对已发表的昆虫MT氨基酸序列进行生物信息学分析。通过分析发现昆虫MT结构保守,序列较短,同一物种不同MT之间同源性低,分类系统不够完善。讨论了昆虫MT研究的难点和不足,并展望了昆虫MT解毒的应用前景。
金属硫蛋白;表达调控;分子特性
1957年,美国学者Margoshes等[1]首次从马肾皮质中分离出一种镉蛋白;随后,Kägi等[2]再从马肾皮质中分离和鉴定一种蛋白质,其每克干质量蛋白质包含镉2.9%、锌0.6%和硫4.1%,并将其命名为“metallothionein”(MT,金属硫蛋白)。之后,MT相继从其它动物和人的肾脏及其它组织中分离出来,而MT的分子特性已成为近十多年来的研究热点。
昆虫是自然界种类最多、数量最大、分布最广的动物,最容易接触到重金属。对昆虫MT的研究能更深入地了解昆虫在受到重金属胁迫下的响应机制,为研究其它动物对重金属的解毒和排毒作用提供参考。
1 MT概述
1.1 MT的分类
MT一般可根据其一级结构分为三大类:Ⅰ型、Ⅱ型和Ⅲ型MT[3]。Ⅰ型MT包括哺乳动物的各种MT,根据其结构和功能的不同,可分为MT-1、MT-2、MT-3和MT-4四大亚型。Ⅱ型MT中半胱氨酸(Cys)的位置与Ⅰ型MT有相似之处,但序列差异较大。Ⅲ型MT主要存在于植物中,是一些具有类似Ⅰ型金属硫蛋白的含Cys序列结构的化合物和多肽。随着更多MT序列的获得,第四届国际金属硫蛋白大会上研究者们提出新的分类系统[2]。将MT定义为一个超家族(Superfamily),并引入家族(Family),再进一步划分为亚家族(Subfamily)、亚群(Subgroup)和异构型(Isoforms)。其中I类和II类MT进一步被分成脊椎动物、软体动物、甲壳动物、棘皮动物、昆虫、线虫、纤毛虫、真菌Ⅰ-Ⅵ、原核生物和植物等15个家族,第Ⅲ类特殊的MT则单独列为99家族(Family 99)[4-5]。
1.2 MT的生理功能
1.2.1 清除自由基 MT清除自由基的功能使其在抗衰老、抗氧化压力及细胞凋亡等过程中发挥着重要作用。MT通过清除多余的自由基,发挥抗氧化、抗辐射作用。研究发现,MT对接受紫外线辐射的晶状体上皮细胞有保护作用[6]。
1.2.2 参与体内微量元素的储存、运输和代谢 在金属元素的代谢中,MT具有维护生物体内金属离子相对稳定的作用。MT能调节小肠对Cu和Zn等金属的吸收,对Zn和Cu代谢起着细胞和细胞内的控制作用[7]。
1.2.3 重金属解毒 MT是富含Cys的金属结合蛋白,其巯基能螯合重金属形成金属复合物,并将之排出体外,从而实现解毒功能[8]。但是MT对重金属解毒和调节作用是有限的,一旦环境中重金属元素含量超过这个阈值,该物种就会出现病理状态。MT是目前临床上最理想的生物螯合解毒剂。
1.2.4 参与应激反应 MT可在应激保护方面发挥作用,如低温和高温、药物中毒、创伤、细菌感染等,都可促进MT的合成。研究证实,MT能提高应激生物体的抗氧化酶的活性,从而降低应激对细胞的损伤作用[9]。
1.3 金属离子对MT的调控
重金属镉可诱导MT的转录表达,其主要调控过程是:镉与MT转录调控因子(metal-responsive transcription factor,MTF-1)结合的转录因子结合,转录调控因子识别MT启动子区金属反应原件(metal response element,MRE)并与细胞核内DNA结合,启动基因的转录[10]。同时,镉胁迫促使生物体产生氧化应激反应,介导MT的表达,此过程可能与MRE有协同作用[11]。重金属镉诱导MT的转录表达,MT大量合成,并与重金属镉形成Cd-MT复合物,同时通过自身疏基还原态氧化态的转换来清除由镉诱导产生的活性氧化基(reactive oxygen species,ROS),减轻镉对生物体的损伤。
2 哺乳动物的MT
哺乳动物的MT-1和MT-2广泛存在于所有动物组织细胞中;MT-3主要在动物神经细胞中表达;MT-4主要在皮肤和上呼吸道表皮细胞中表达[12-14]。典型的哺乳动物MT分子量为6 000~7 000 Da,由60~68个氨基酸残基组成,其中Cys含量为20%~30%,且位置非常保守[15]。MT与金属离子结合形成两个大小相当的结构域和,彼此单独呈球形,中间通过氨基酸残基连接,使整个分子呈哑铃状[16]。
3 植物的MT
植物的MT在根、茎、叶、花、果实和种子中的表达存在组织特异性[17]。Ⅰ亚型MT在根中表达比较丰富,Ⅱ亚型主要表达在叶中,Ⅲ亚型MT的RNAs在成熟的肉质果或叶中高表达,Ⅳ亚型MT在发育着的种子中表达受到抑制[18-20]。植物MT蛋白两端富含Cys的区域相互接近和金属离子结合形成一个结构域。目前植物MT有hairpin、Dumbbell和double dumbbell等3种结合模型[21]。
4 昆虫的MT
4.1 昆虫MT的分布
昆虫体内的MT主要存在于消化道,尤其在中肠。经解剖学研究发现,大约90%的Cd存在于麻蝇幼虫()的消化道[22],而这些Cd主要与蛋白质结合以MT的形式存在。通过109Cd2+放射性原子示踪试验,检测到黑腹果蝇()幼虫摄入体内的95%以上放射性物质分布在幼虫中肠[23]。
4.2 昆虫MT的结构
目前,已知昆虫的MT分子量为5 000 Da左右,编码45个左右氨基酸,部分昆虫中含有1%~2%的芳香族氨基酸和组氨酸[24]。果蝇金属硫蛋白的2个基因MTA和MTB分别编码40和43个氨基酸,其中仅11个氨基酸残基是相同的,只有25%的同源性,这显然不同于哺乳动物[25]。
4.3 昆虫MT的功能
4.3.1 重金属解毒 重金属可诱导昆虫MT合成,从而保护机体免受重金属毒害。中华稻蝗()基因沉默后,昆虫对重金属的耐受性明显下降,表明金属硫蛋白可提高昆虫对重金属的耐受性[26]。家蝇()幼虫在不同浓度的镉诱导24 h后,在半致死浓度胁迫下家蝇基因的表达量显著高于对照,说明金属硫蛋白能结合镉并将其排出体外[27]。
4.3.2 生物标志物 当生物体暴露于重金属时,都能很明显地监测到MT的变化,因此MT可作为监测环境重金属污染的标志物。黑水虻()MT的表达量在一定范围内随着镉浓度的升高而升高,表明它在废物处理过程中可能具有作为生物标志物监测镉的潜力,可应用于环境污染检测[28]。
5 昆虫MT的序列分析研究
5.1 已发表的昆虫MT
虽然MT在多种哺乳动物中得到了分离和鉴定,但昆虫基因的研究仅在少数种类中有公布,其中以果蝇的MT研究最为深入,其它昆虫中已鉴定的基因序列均不超过3条,如近期进行序列鉴定的直翅目昆虫中华稻蝗。已经发表的昆虫MT序列信息见表1(属以黑腹果蝇为代表)。金属硫蛋白序列下载自GenBank(http://www.ncbi.nlm.nih.gov/genbank/)。
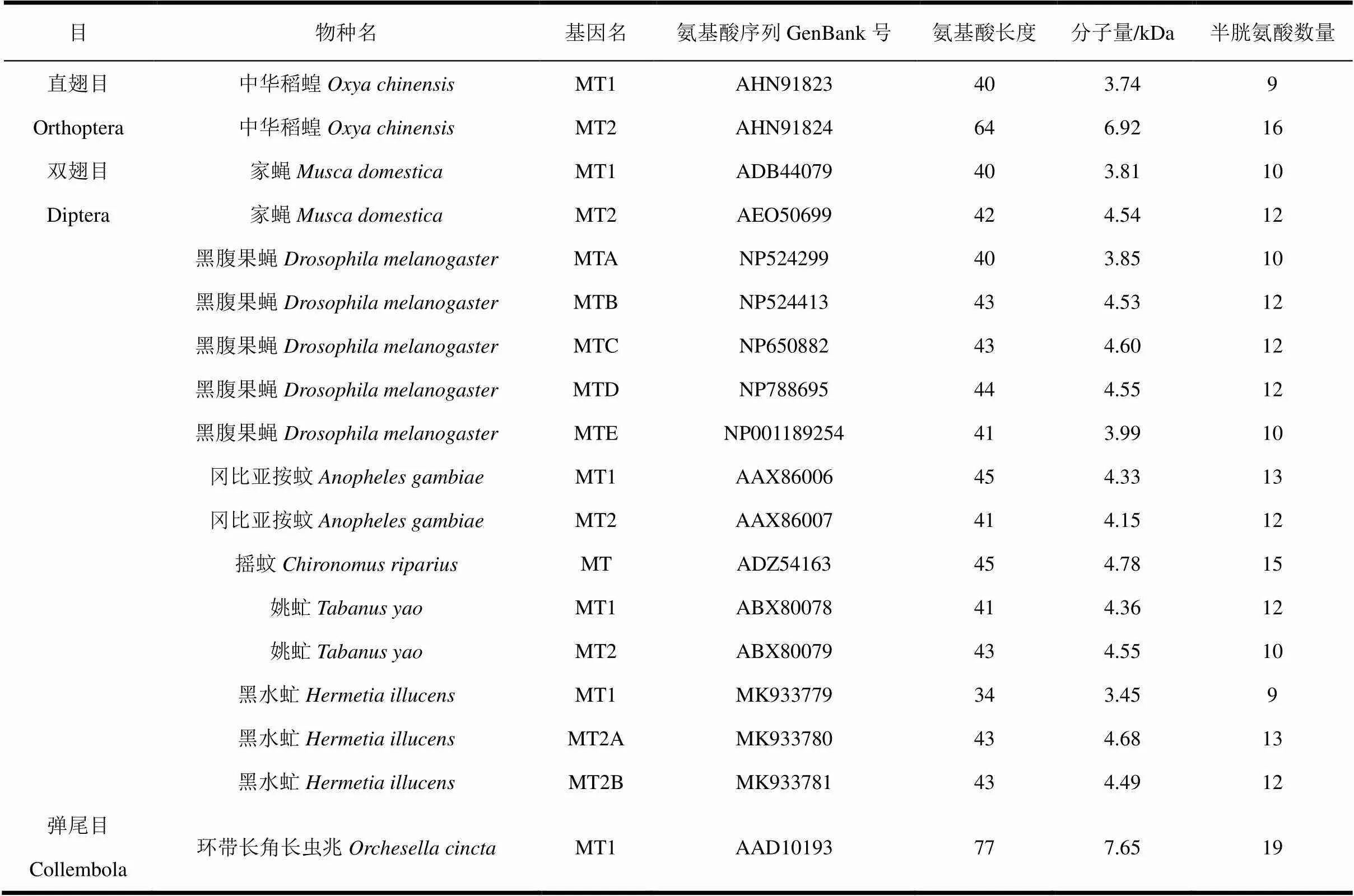
表1 已发表的昆虫MTs
5.2 昆虫MT的氨基酸序列比对分析
采用Mega 6.0和Genedoc软件对这些昆虫的氨基酸序列进行比对,这7种昆虫Cys含量为25%左右(图1),大多数位于保守位点上,说明MT蛋白在昆虫中高度保守。通过7种昆虫的MT对比发现,不同物种的金属硫蛋白在氨基酸数量及组成等方面存在差异,Cys的分布和排列方式是非常保守的,多为-C-X-C-(C为半光氨酸,X是除半胱氨酸以外的其他氨基酸),说明Cys对金属硫蛋白的功能极其重要。

黑腹果蝇Drosophila melanogaster (DmMTA, NP524299; DmMTB, NP524413; DmMTC, NP650882; DmMTD, NP788695; DmMTE, NP001189254); 中华稻蝗Oxya chinensis (OcMT1, AHN91823; OcMT2, AHN91824); 家蝇Musca domestica (MdMT1, ADB44079; MdMT2, AEO50699); 冈比亚按蚊Anopheles gambiae (AgMT1, AAX86006; AgMT2, AAX86007); 摇蚊Chironomus riparius (CrMT, ADZ54163); 姚虻Tabanus yao (TyMT1, ABX80078; TyMT2, ABX80079); 黑水虻Hermetia illucens (HiMT1, MK933779; HiMT2A, MK933780; HiMT2B, MK933781); 环带长角长虫兆Orchesella cincta (OcMT1, AAD10193)。
5.3 昆虫MTs的系统发育树分析
采用Mega 6.0软件基于NJ法建立MT分子系统发育树(图2),从进化树上可以看出,黑水虻MT1,黑腹果蝇MTA,中华稻蝗MT1和家蝇MT1聚在一起。中华稻蝗的两个基因的进化关系较远,黑腹果蝇中的MTB、MTC、MTD和MTE的进化关系较近。

图2 昆虫MTs的系统发育树分析
6 小结与展望
MT作用广泛,其重金属解毒等生物活性功能越来越多的被开发出来。例如,MT可作为生物标志物,水生无脊椎动物和鱼暴露于重金属中时,能很明显地监测到MT的变化[29],能有效地表征水体中重金属对生物的影响,多应用于环境污染研究;MT还应用于治疗疾病、作为向导药物载体开发生物药剂等,实验证明心肌缺血再灌注损伤时,金属硫蛋白2A能发挥保护心肌的作用,从临床研究、动物实验和体外实验等多个层面揭示金属硫蛋白2A在心脑血管疾病中的可能作用[30]。
本文根据序列同源性和进化关系研究昆虫MT的分子特征,这种特征研究是基于已有序列归纳的特征,因此,合理的特征分类体系的建立依赖于更多MT序列的获得。目前已知种类和数量有限,昆虫基因序列较短不易克隆;另一方面,MT蛋白分离纯化后,其自然活性不易维持,而且可能存在重金属污染,不适合用于大规模的产品开发。昆虫MT的研究领域主要集中在MT的分离纯化、理化特性等方面,对昆虫MT功能和体外表达方面的研究并不全面。因此,今后的研究中应整合昆虫的生物学相关技术和方法,从基因、蛋白、以及体外表达方面进行研究,为昆虫MT的表达调控和相互作用机制的进一步研究提高理论与实验依据,最终实现稳定的体外表达并应用到实际中。
[1] MARGOSHES M, VALLEE B L. A cadmium protein from equine kidney cortex.[J]. Journal of the American chemical society, 1957, 79(17): 4813-4814.
[2] KÄGI J H R, VALLEE B L. Metallothionein: a cadmium-and zinc-containing protein from equine renal cortex.[J]. The journal of biological chemistry, 1960, 235(9): 3460.
[3] EBADI M, IVERSEN P L, HAO R, et al. Expression and regulation of brain metallothionein[J]. Neurochemistry international, 1995, 27(1): 1-22.
[4] BINZ P A , JHR K. Metallothionein IV[M]. Switzerland: Library of Congress Cataloging-in-publication Data, 1999: 3-6.
[5] KOJIMA Y, BINZ P A, JEREMIAS H R. Metallothionein IV [M]. Switzerland: Library of Congress Cataloging-in-publication Data, 1999: 7-14.
[6] SAITO T, TEZUKA T, KONNO R, et al. Protective effects of metallothionein I and II against metal- and ultraviolet radiation-induced damage in cultured lens epithelial cells[J]. Japanese journal of ophthalmology, 2010, 54(5): 486-493.
[7] 茹炳根, 潘爱华, 黄秉乾, 等. 金属硫蛋白[J]. 生物化学与生物物理进展, 1991(4): 12-17.
[8] 路浩, 刘宗平, 赵宝玉. 金属硫蛋白生物学功能研究进展[J]. 动物医学进展, 2009, 30(1): 62-65.
[9] 刘湘新, 李丽立, 刘进辉, 等. 金属硫蛋白对应激猪血清中某些酶活性的影响[J]. 中国兽医杂志, 2005, 41(12): 8-9.
[10] RADTKE F, HEUCHEL R, GEORGIEV O, et al. Cloned transcription factor MTF-1 activates the mouse metallothionein I promoter[J]. The embo journal, 1993, 12(4): 1355-1362.
[11] ANDREWS G K. Regulation of metallothionein gene expression by oxidative stress and metal ions[J]. Biochemical pharmacology, 2000, 59(1): 95-104.
[12] STILLMAN M J. Metallothioneins[J]. Coordination chemistry reviews, 1995, 144(10): 461-511.
[13] TOKUDA E, ONO S I, ISHIGE K, et al. Metallothionein proteins expression, copper and zinc concentrations, and lipid peroxidation level in a rodent model for amyotrophic lateral sclerosis[J]. Toxicology, 2007, 229(1/2): 33-41.
[14] TAKAHASHI T, GAO X D. Physical interactions among human glycosyltransferases involved in dolichol-linked oligosaccharide biosynthesis[J]. Trends in glycoence & glycotechnology, 2012, 24(136): 65-77.
[15] 韩新燕, 许梓荣. 哺乳动物金属硫蛋白的研究进展[J]. 中国兽医科学, 2003, 33(11): 28-32.
[16] NIELSEN A E, BOHR A, PENKOWA M. The Balance between life and death of cells: roles of metallothioneins[J]. Biomarker insights, 2006, 1(1): 99-111.
[17] 孟红恩, 刘忠渊. 植物金属硫蛋白研究进展[J]. 广东农业科学, 2014, 41(15): 133-138.
[18] YANG Z, WU Y, LI Y, et al. OsMT1a, a type 1 metallothionein, plays the pivotal role in zinc home ostasis and drought tolerance in rice[J]. Plant molecular biology, 2009, 70(1/2): 219-229.
[19] HUANG G Y, WANG Y S. Expression analysis of type 2 metallothionein gene in mangrove species () under heavy metal stress[J]. Chemosphere, 2009, 77(7): 1026-1029.
[20] HUANG G Y, WANG Y S, YING G G. Cadmium-inducible BgMT2, a type 2 metallothionein gene from mangrove species () , its encoding protein shows metalbinding ability[J]. Journal of experimental marine biology and ecology, 2011, 405(1/2): 128-132.
[21] 翟军鹏, 夏蓓蓓, 田婷婷, 等. 植物金属硫蛋白的研究进展[J]. 南方农业, 2010, 34(1): 87-89.
[22] AOKI Y, SUZUKI K T, KUBOTA K. Accumulation of cadmium and induction of its binding protein in the digestive tract of fleshfly () larvae[J]. Comparative biochemistry and physiology part C: comparative pharmacology, 1984, 77(2): 279-282.
[23] MARONI G, WATSON D. Uptake and binding of cadmium, copper and zinc bylarvae[J]. Insect biochemistry, 1985, 15(1): 55-63.
[24] HENSBERGEN P J, VELZEN M J M V, NUGROHO R A, et al. Metallothionein-bound cadmium in the gut of the insect(Collembola) in relation to dietary cadmium exposure[J]. Comp biochem physiol c toxicol pharmacol, 2000, 125(1): 1-24.
[25] 牛长缨, 姜勇, 雷朝亮, 等.无脊椎动物金属硫蛋白的研究[J]. 动物学杂志, 2002, 37(1): 72-76.
[26] 张迪, 任国栋, 唐婷,等. 家蝇金属硫蛋白基因的克隆、原核表达及活性检测[J]. 昆虫学报, 2010, 53(4): 379-384..
[27] 刘耀明, 余志涛, 朱文雅, 等. 三种重金属对中华稻蝗金属硫蛋白基因表达的影响[J]. 农业环境科学学报, 2015, 34(2): 227-232.
[28] MOURGAUD Y, MARTINEZ, E, GEFFARD A, et al. Metallothionein concentration in the mussel Mytilus galloprovincialis as a biomarker of response to metal contamination: validation in the field[J]. Biomarkers, 2002, 7(6): 479-490.
[29] WANG X, GAO Q, LIU X, et al. Metallothionein in(Linnaeus, 1758) larvae (Diptera: Stratiomyidae), a potential biomarker for organic waste system[J]. Environmental science and pollution research, 2018, 25(6): 5379-5385.
[30] 戴明强, 焦蓉, 徐彩霞, 等. 金属硫蛋白2A与心脑血管疾病的研究进展[J]. 中国动脉硬化杂志, 2018, 26(3): 310-314.
Research Progress of Insect Metallothionein
ZHANG Jiali, ZHOU Xiaocao, LUO Mei, WEI Hongyi*
(School of Agronomy Science, Jiangxi Agricultural University, Nanchang 330045, China)
Metallothioneins (MT) play roles in scavenging free radicals, regulating the concentration of trace elements in organisms, and participating in the detoxification of heavy metals. This article introduced the biological characteristics, expression regulations and physiological functions of MT, and conducted bioinformatic analysis of the published amino acid sequence of insect MT. It was found that insect MT structure was conservative, but the sequence was short, the homology between different MTs of the same species was low, and the classification system was not perfect. The article also discussed the difficulties and shortcomings of insect MT researches, and looked forward to the application prospects of insect MT detoxification.
metallothionein; expression regulation; molecular characteristics
Q965
A
2095-3704(2021)04-0391-06
张佳丽, 周小草, 罗妹, 等. 昆虫金属硫蛋白的研究进展[J]. 生物灾害科学, 2021, 44(4): 391-396.
2021-11-06
2021-12-13
国家自然科学基金项目(31760637,31640064)
张佳丽(1996—),女,硕士生, 主要从事农业昆虫与害虫防治研究,Zhangjiali622@outlook.com;
通信作者:魏洪义,教授,hywei@jxau.edu.cn。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
大自然探索(2023年11期)2023-03-01 09:04:36
玩具世界(2022年3期)2022-09-20 01:48:20
学苑创造·A版(2022年3期)2022-03-29 23:32:16
昆明医科大学学报(2021年3期)2021-07-22 07:40:08
甘肃教育(2020年2期)2020-09-11 08:01:48
制造技术与机床(2019年10期)2019-10-26 02:47:12
学苑创造·A版(2019年6期)2019-07-11 01:07:39
小学生优秀作文(低年级)(2018年9期)2018-09-10 05:47:38
小学生必读(低年级版)(2018年11期)2018-03-13 00:54:24
