
车前种子黏液对其劣变和萌发抗逆性的影响
2022-01-08 02:54叶晓倩刘瑞曦冯金慧邓仕明李吉涛邓志军
草地学报 2021年12期
叶晓倩, 刘瑞曦, 冯金慧, 邓仕明, 李吉涛, 邓志军
(1. 生物资源保护与利用湖北省重点实验室, 湖北民族大学, 湖北 恩施 445000; 2. 恩施州特色植物资源种质工程技术研究中心, 湖北民族大学, 湖北 恩施 445000; 3. 湖北民族大学林学园艺学院, 湖北 恩施 445000)
种子黏液是由种皮外层细胞中的高尔基体产生并分泌到细胞腔或细胞壁的一些具有强亲水性的酸性(果胶)和中性(阿拉伯糖、纤维素、甘露糖、鼠李糖和木糖)多糖[1-2]。研究发现,至少37目110科230属的被子植物的种子具有黏液[3-4]。种子黏液可以促进种子成熟和传播,可以通过保湿并激活种子的修复机制来保持种子库的有效性,可以通过减少氧扩散来维持种子休眠,可以通过调节水分和氧气的吸收促进种子萌发,还可以通过直接或间接提供营养促进幼苗生长[5]。尤其是在沙漠条件下,种子黏液使很多荒漠植物在干旱少雨条件下能够及时快速萌发成苗[6]。
在自然条件下,种子最终能否萌发成苗,既取决于种子活力的保持能力,也取决于种子的萌发抗逆性。目前,已知种子黏液有利于种子传播与萌发成苗,特别是有利于种子在干旱等逆境条件下的萌发成苗,而种子黏液对种子活力保持的影响,以及种子黏液在种子活力保持与萌发抗逆性之间的关系中的作用还尚未明确。对这方面的研究将有助于进一步全面认识种子黏液的生态意义。
车前(Plantagoasiatica)为车前科(Plantaginaceae)车前属多年生草本植物,全国广泛分布[7]。车前种子及全草皆可入药[8],且在园林绿化中也有广阔应用前景[9]。车前和同属的小车前(P.minuta)[10]、长叶车前(P.lanceolata)[11]、平车前(P.depressa)[12]、盐生车前(P.maritimasubsp.Ciliata)[13]和北美车前(P.virginica)[14]的种子表面均具有黏液,这似乎是车前属植物的共有特征。目前已知,北美车前[14]和小车前[15]种子的黏液均有利于其在干旱环境下萌发成苗,而种子黏液对车前种子萌发抗逆性和活力保持的影响则未见报道。
1 材料与方法
1.1 试验材料
车前种子于2018年6月采自湖北省鹤峰县木林子国家级自然保护区。采回的种子在室温下阴干一周,测定含水量((8.36±0.02)%)后装入聚乙烯密封袋并置于4℃冰箱中保存备用。
1.2 种子黏液去除
将种子平铺在用蒸馏水充分湿润的双层滤纸上,在10℃黑暗条件下培养4 h,使种子表面黏液物质充分吸水软化,然后将吸胀后的种子裹在四层医用纱布间用手轻轻揉搓以使黏液物质与种子充分分离,再通过蒸馏水清洗去除黏液,最后平铺在干燥滤纸上于10℃黑暗条件下阴干,直至含水量降至与初始含水量相近水平。
1.3 人工加速老化处理
参照Liu等[16]的方法将保留和去除黏液的车前种子分别置于40℃和100%相对湿度下进行人工加速老化处理。在老化处理0,2,4,6,8,10,12,14,16,18 d后取样,并在指定的最适温度下进行萌发测试。
1.4 模拟干旱处理
参照Michael和Kaufaman[17]的方法,配制水势分别为0,-3,-4,-5,-7,-8,-9,-10 MPa的PEG-6000溶液来模拟不同程度的干旱条件,然后用这些不同水势的PEG-6000溶液在指定的最适温度下同步培养保留和去除黏液的车前种子。
1.5 模拟盐胁迫处理
用0,50,75,100,125,150,175,200 mmol·L-1的NaCl溶液来模拟不同程度的盐胁迫条件,然后用这些不同浓度的NaC l溶液在指定的最适温度下培养保留和去除黏液的车前种子。
1.6 萌发测试
新采收的种子分别在5,10,15,20,25,30,35,40℃恒温,以及15℃/25℃和20℃/30℃变温(每天低温和高温各12 h)和交替光照(12 h黑暗/12 h光照,光照强度为121 μmol·m-2·s-1,变温条件下的低温与高温时间段分别对应于黑暗与光照时间段)条件下测试初始萌发,以选定种子萌发的最适温度。
去除黏液和保留黏液种子经人工加速老化处理、模拟干旱处理、盐胁迫处理后在选定的最适恒温下进行萌发测试。每个处理设4次重复,每次重复50粒种子,以胚根突破种皮2 mm作为萌发完成的标准,连续5 d不再有新增加的萌发种子后结束萌发测试,统计最终萌发率。萌发速率以从培养开始到萌发率达最终萌发率的50%时所用的时间(T50)表示,T50值与萌发速率成反比,即T50值越小,萌发速率越大。具体计算方法是,先对种子累积萌发率随培养时间的变化进行曲线回归,然后通过回归方程计算出T50值。通过下式计算种子萌发率:
种子萌发率(%)=萌发的种子数/供试种子数×100
1.7 数据处理与分析
统计分析和作图均使用R软件(R i386 3.5.2)完成。所有的萌发率数据和T50数据均采用单因素方差分析(One-way ANOVA,P=0.05),并进行Student-Newman-Keuls验后多重比较(P=0.05)。文中所有统计数据均以“平均值±标准误”形式表示。
2 结果与分析
2.1 温度对新鲜成熟车前种子萌发的影响
如图1所示,温度对新鲜成熟车前种子萌发具有显著影响(P<0.05)。车前种子除了在5℃和40℃恒温条件下无萌发外,在其他温度下均有不同数量的萌发。在25℃与30℃恒温及15℃/25℃与20℃/30℃变温条件下的萌发率间无显著差异,均在94%以上,且均显著高于其他温度条件下的萌发率。选定25℃作为人工加速老化处理、模拟干旱处理、盐胁迫处理后的萌发测试温度。
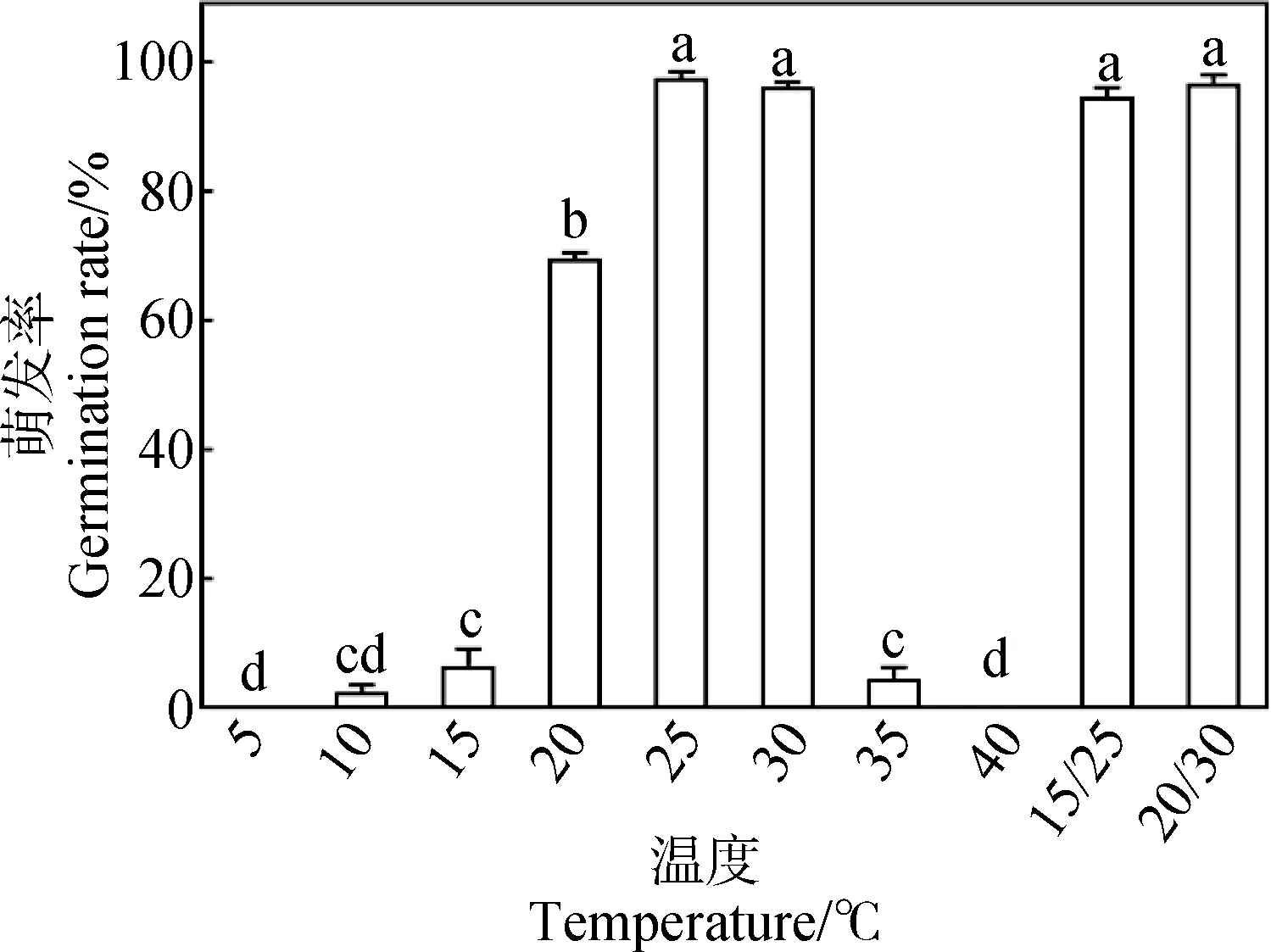
图1 新鲜成熟车前种子在几个恒温、交替变温和交替光照条件下的萌发率
2.2 去除黏液和保留黏液种子萌发对人工加速老化的响应
随着老化处理时间的延长,去除黏液和保留黏液种子的萌发率(图2A)和萌发速率(图2B,萌发速率与T50值成反比)均呈逐渐下降趋势,保留黏液种子老化8 d后萌发率即显著下降,而去除黏液种子则直到老化12 d后萌发率才发生显著下降,两种种子均在老化18 d后几乎不再有种子萌发(图2A)。经6 d的老化处理后,去除黏液和保留黏液种子的萌发率无显著差异(图2A),但去除黏液种子的萌发速率却显著高于保留黏液种子(P<0.05,图2B);经10 d和12 d的老化处理后,去除黏液种子的萌发率和萌发速率均显著高于保留黏液种子(P<0.05,图2A,B);经14 d的老化处理后,去除黏液种子的萌发率显著高于保留黏液种子(P<0.05,图2A),但两者的萌发速率间却无显著差异(图2B);对于其他老化处理,两种种子在萌发率和萌发速率方面均无显著差异(图2A,B)。
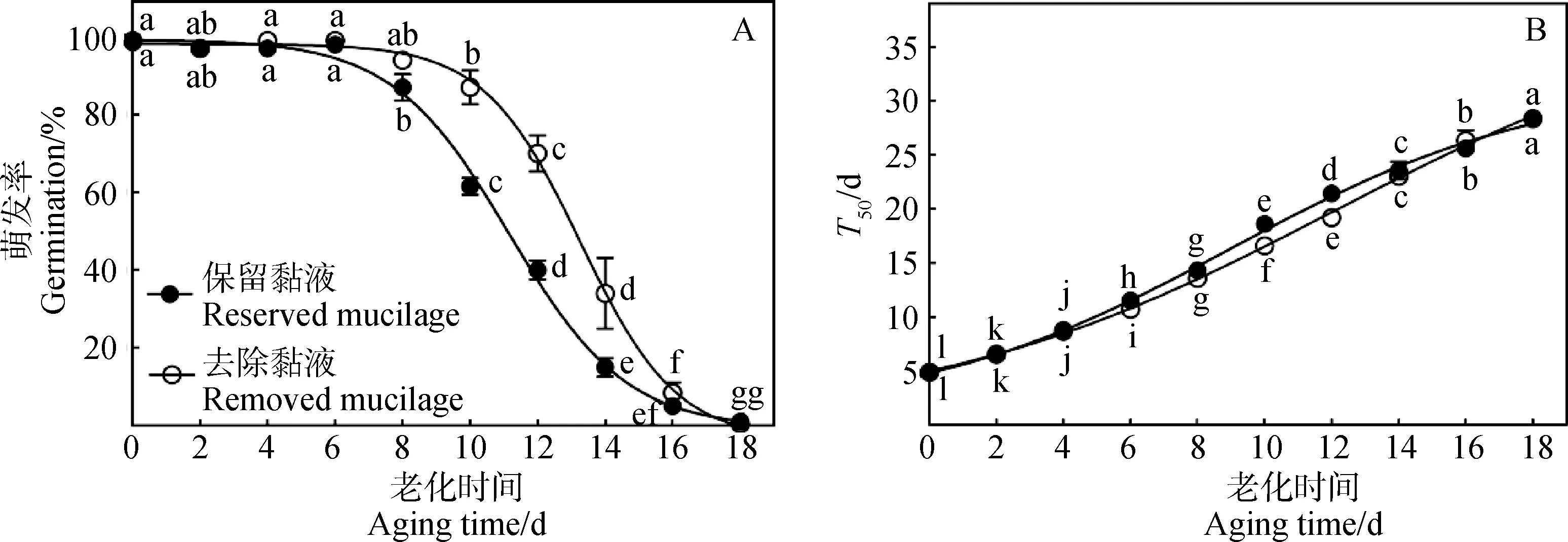
图2 车前种子的萌发率(A)和萌发速率(B)随人工加速老化处理的变化
2.3 去除黏液和保留黏液种子萌发对干旱胁迫的响应
随着PEG-6000溶液水势的降低(即干旱胁迫增强),不管种子有无黏液,车前种子的萌发率(图3A)和萌发速率(图3B)均呈下降趋势。所不同的是,保留黏液种子的萌发率在水势降至-5 MPa时开始显著下降,随后则相对缓慢地下降;而去除黏液种子萌发率则当水势降至-7 MPa时才显著下降,但随后却相对更加快速地下降,当水势小于等于-9 MPa时均不再有种子萌发(图3A)。在-5 MPa水势条件下去除黏液种子的萌发率显著高于保留黏液种子(P<0.05),而在-7 MPa水势条件下保留黏液种子的萌发率则显著高于去除黏液种子(P<0.05),在其他水势条件下两者的萌发率均无显著差异(图3A);而从萌发速率来看,除了在-8 MPa水势条件下保留黏液种子的萌发速率显著高于去除黏液种子外(P<0.05),在其他水势条件下两者均无显著差异(图3B)。

图3 车前种子萌发率(A)和萌发速率(B)随干旱胁迫处理的变化
2.4 去除黏液和保留黏液种子萌发对盐胁迫的响应
随着NaCl溶液浓度的升高(盐胁迫增强),去除黏液和保留黏液种子的萌发率(图4A)和萌发速率(图4B)均具有相似的变化趋势。均当NaCl溶液浓度升高至175 mmol·L-1的时候,萌发率才急剧下降,而当NaCl溶液浓度升高至200 mmol·L-1时种子均几乎不再萌发(图4A)。从萌发速率来看,仅在75 mmol·L-1的NaCl溶液培养下保留黏液种子的萌发率速率显著高于去除黏液种子(P<0.05),其余各浓度NaCl溶液处理下两种种子的萌发速率均无显著差异(图4B)。

图4 车前种子萌发率(A)和萌发速率(B)随盐胁迫处理的变化
3 讨论
种子劣变是一种自然现象,同时受到自身遗传因素和外界环境条件的影响。本研究通过人工老化处理模拟自然劣变,从而探讨种子黏液对种子活力保持的作用。结果显示,随着老化程度的加深,去除黏液种子的萌发速率和萌发率几乎均高于保留黏液种子,说明黏液物质的存在不利于种子生活力的保持。已有的研究表明,保留黏液种子的吸水量显著高于去除黏液种子的吸水量[14-15]。因此,在人工老化(高温高湿)条件下,保留黏液的车前种子的吸水量肯定高于去除黏液种子,使保留黏液种子的呼吸作用更强,从而加速种子内贮藏物质的消耗,最终导致生活力丧失。同时,呼吸作用的增强会放出更多的热量,进而导致种子表面及种子间的微生物活动增强,从而产生更多的热量,最终致使种子发霉变质。
对于干旱环境下的种子而言,水分是其萌发成苗的决定因子[18]。种子黏液可通过调节水分吸收来调控种子萌发,以适应多变的水分环境,从而增强种子的抗旱性[5]。伍晨曦等[15]发现,在低渗透胁迫(-0.33~-0.15 MPa)下,小车前保留黏液种子的发芽势和萌发率均显著高于去除黏液种子;而在高渗透胁迫(-1.15~-0.73 MPa)下,种子黏液的有无对种子萌发率无显著影响。罗辉等[14]则发现,仅在高渗透胁迫(-1.21~-0.58 MPa)下,保留黏液的北美车前种子的萌发率显著高于去除黏液种子。但在本研究中,随着渗透胁迫的加剧,种子黏液对于车前种子萌发抗旱性的影响,在萌发率和萌发速率两个方面具有不同的表现,在-4~0 MPa水势条件下,两种种子在萌发率和萌发速率上均无显著差异;在相对较低渗透胁迫(-5 MPa)下时,保留黏液种子的萌发率显著低于去除黏液种子;在相对中等水平的渗透胁迫(-7 MPa)下时,保留黏液种子的萌发率显著高于去除黏液种子;而在相对较高渗透胁迫(-8 MPa)下时,两种种子的萌发率无显著差异,只是保留黏液种子的萌发速率显著高于去除黏液种子。由此可以推断,萌发速率比萌发率对渗透胁迫的响应更为敏感,种子黏液在较低渗透胁迫下似乎阻碍种子萌发,仅在较高渗透胁迫下才表现出促进萌发的作用。
种子萌发期对盐敏感,盐胁迫会抑制种子萌发[19]。车前种子[20]、小车前种子[21]的萌发率均随着盐溶液浓度的升高而降低。在本研究中,车前种子的萌发率随着盐浓度的升高大致呈下降趋势,直至几乎不能萌发。从萌发率来看,在相同盐浓度下保留或去除黏液对车前种子的萌发率没有显著影响,从萌发速率来看,仅在75 mmol·L-1的NaCl溶液培养下保留黏液种子的萌发速率显著高于去除黏液种子。由此可见,从对萌发速率的影响来看,车前种子黏液可增强种子对轻微盐胁迫的萌发抗性。盐胁迫对植物的危害常表现为水分胁迫,外界与细胞间不当的渗透势差迫使细胞失水,严重时甚至发生质壁分离、细胞死亡等现象[22]。种子黏液通过调节水分,消除不当渗透差,防止种子过度失水,从而达到减缓盐害的目的[23]。可以看出,车前种子的黏液对于种子在干旱和盐胁迫下的萌发具有一定的调节作用,在较低程度的干旱和盐胁迫条件下,黏液物质的存在保证种子表面有足够的有效水分,减缓了干旱和盐胁迫对种子萌发的不利影响,从而保障了种子正常吸水萌发。
4 结论
车前种子黏液物质的存在,一方面,不利于种子活力的保持,加速其老化,而另一方面,又在一定程度上提高了种子对轻微干旱胁迫和盐胁迫的抗性。
猜你喜欢
当代水产(2022年6期)2022-06-29
干旱地区农业研究(2021年6期)2021-12-08
农村实用技术(2020年9期)2020-11-19
中国临床医学影像杂志(2019年5期)2019-08-27
中国临床医学影像杂志(2019年2期)2019-04-25
现代检验医学杂志(2016年3期)2016-11-15
中国马铃薯(2016年3期)2016-09-07
植物资源与环境学报(2016年1期)2016-08-12
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
节水灌溉(2016年3期)2016-03-23
