
蓟马为害对苜蓿叶茎显微结构的影响
2022-01-08 08:13温雅洁周生英方强恩胡桂馨
草地学报 2021年12期
温雅洁, 周生英, 方强恩, 胡桂馨
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 中-美草地畜牧业可持续发展中心, 甘肃 兰州 730070)
在长期的进化历程中,植物和昆虫形成了一个既对立又统一的体系,寄主植物为植食性昆虫提供食物和栖息场所的同时,形成了自我保卫机制以抵御或适应昆虫取食为害[1]。植物叶片表面的茸毛密度和长短会影响昆虫的寄生、采食和繁殖,叶片较厚的蜡质会增强叶片对光波的反射能力,影响昆虫对寄主的准确定位,叶片上气孔的排列疏密、大小也影响昆虫采食,表明植物叶片表皮的特殊结构会增强其抗虫性[2]。植物的形态结构与生理功能具有一致性,研究表明植物叶片解剖结构和光合器官解剖学特征与叶片光合速率紧密相关[3-4]。叶绿体和基粒数量的增加,能更有效地截获光能,进而提高作物净光合速率和产量[5]。因此,了解植物组织结构与昆虫取食胁迫的关系,不仅有助于进一步探索植物与昆虫的相互适应关系,而且对利用植物抗虫性防治害虫、提高作物产量也具有重要的意义。
由于蓟马发育繁殖速度快、繁殖力强和世代重叠严重等特性,致使苜蓿在第二、三茬的受害率达到100%,苜蓿干草损失量达14%以上[6]。以牛角花齿蓟马(Odontothripsloti)为优势种的蓟马类害虫的危害已成为我国西北、华北和东北等苜蓿主产区的重要威胁和障碍[6-7]。胡桂馨等[8-9]和贺春贵等[10]通过室内和大田抗蓟马评价,揭示了苜蓿对蓟马的抗虫机制主要表现为耐害性。蓟马为害影响了苜蓿植株的物理性状,受蓟马为害后苜蓿茎秆变细,茎秆与叶片颜色加深,尤其是抗蓟马苜蓿叶片颜色变化更大[11]。寇江涛等[12]的研究发现,紫花苜蓿对蓟马的危害具有明显的光合补偿作用,受害后抗蓟马苜蓿无性系R-1光合补偿效应显著大于感蓟马苜蓿无性系I-1。但受蓟马为害后苜蓿组织显微和超显微结构的变化尚不清楚。本试验选取抗蓟马苜蓿无性系R-1和感蓟马苜蓿无性系I-1为材料,观测苜蓿大田蓟马为害后抗、感蓟马苜蓿无性系的叶片、茎秆组织显微结构及叶片超微结构的变化,探索苜蓿组织结构与抗蓟马特性间的联系,为苜蓿抗蓟马研究提供理论依据。
1 材料与方法
1.1 试验材料
供试验用苜蓿材料:抗蓟马苜蓿无性系R-1、感蓟马无性系I-1。
供试昆虫:以牛角花齿蓟马(Odontothripsloti)为优势种的苜蓿大田蓟马复合种群。
1.2 试验设计
试验在甘肃农业大学草业学院牧草试验站中进行。4月上旬,将室内扦插的抗蓟马苜蓿无性系R-1和感苜蓿无性系I-1植株移栽于大田老苜蓿地附近,抗、感苜蓿隔行栽植,行距30 cm,株距20 cm,每种无性系材料各3行,每行30株,试验地面积12 m2。6月下旬,第1茬刈割后,用塑料布将试验地隔为两半,一半试验地每15 d喷洒一次吡虫啉控制,另一半试验地任由蓟马为害至苜蓿现蕾前期(现蕾率达到20%),取样观察抗、感蓟马苜蓿叶片和茎秆组织结构的变化。
1.3 观测指标与试验方法
1.3.1抗、感苜蓿健康植株与受害植株叶片上、下表皮的变化 每行随机选3株植株,选取健康植株倒5叶和受害植株上受害级别为2级[10]的倒5叶,复叶中间小叶的中部、距主脉两侧5 mm左右部位,受害叶选取叶中部未受害部分。用刀片切取边长2.00~2.50 mm的正方形,立即投入盛有4%戊二醛固定液的小瓶中,于4℃保存。利用扫描电子显微镜(Hitachi-S 3400 N)观测单位面积表皮细胞和气孔数量,每处理每无性系共随机观测30个视野,取均值。
1.3.2抗、感苜蓿健康植株与受害植株茎秆表皮的变化 每行随机选3株,取健康植株和受害植株茎秆,于第一朵花蕾下第二节基部1 cm处,用宽头镊子剥下表皮,用1%龙胆紫染色,制成玻片,在生物显微镜(Panthera U,麦克奥迪实业集团有限公司)下观测茎秆棱角和平面单位面积表皮细胞和气孔数量,每处理每无性系共随机观测30个视野,取均值。
1.3.3抗、感苜蓿健康植株与受害植株茎秆维管束结构的变化 每行随机选3株,取健康植株和受害植株茎秆,于第一朵花蕾下第二节基部1 cm处,徒手切片,挑取较完整的切片,用固绿染色,制成玻片标本,在生物显微镜下观测苜蓿受害株和健康株的茎秆维管束结构变化,每处理每无性系共随机观测30个视野。
1.3.4抗、感苜蓿健康植株与受害植株叶片叶肉细胞和叶绿体的变化 每行随机选3株,健康植株倒5叶和受害植株级别为2级的倒5叶,选取叶片中部、距主脉5 mm部位的正、反面(受害叶取未受害部分),取边长2.00~2.50 mm的正方形叶片,立即投入盛有4%戊二醛固定液的小瓶中,4℃保存。采用常规电镜制样方法,经四氧化锇双固定、乙醇系列脱水、环氧树脂包埋,超薄切片机切片,柠檬酸铅和乙酸双氧铀双重染色,在JEM-1230型透射电镜(日本电子株式会社广州事务所)下观察和拍照,观察抗、感植株叶片组织结构(包括栅栏组织和海绵组织、叶绿体结构特征),每处理每无性系共随机观测30个视野。
1.4 数据处理
用Microsoft Excel 2010软件进行试验数据处理,制作图表,并用SPSS 20.0软件进行T检验(P<0.05)。
2 结果与分析
2.1 抗、感苜蓿健康植株与受害植株叶片表皮特征的变化
由表1可知,蓟马为害后,单位面积细胞数和气孔数显著增加(P<0.05)。健康抗蓟马苜蓿R-1叶片上、下表皮单位面积细胞数和气孔数较健康感蓟马苜蓿I-1少,但受害后上表皮单位面积细胞数和气孔数R-1均高于I-1,而下表皮单位面积细胞数和气孔数R-1均低于I-1。从增加的倍数来看,受蓟马为害后,R-1的上表皮和下表皮单位面积细胞数的增加倍数分别为1.75和1.51,单位面积气孔数的增加倍数分别为1.67和1.37,均高于I-1。

表1 健康与受害R-1和I-1苜蓿叶片上、下表皮细胞与气孔数目比较
由图1可知,健康抗、感苜蓿叶片的上表皮细胞与下表皮细胞形态特征一致,表皮细胞形状均不规则,大小不一,排列紧密,垂周壁式样为波形,细胞边缘呈锯齿状(图1A,1C,1E,1G)。受蓟马为害后,抗、感苜蓿叶片上下表皮细胞显著隆起、形状变为多边形,垂周壁式样变为平直形,锯齿状边缘消失(图1B,1D,1F,1H)。

图1 R-1和I-1苜蓿健康和受害叶片上、下表皮细胞(500X)
2.2 抗、感苜蓿健康株与受害株叶片显微结构的变化
由图2可知,蓟马为害后,抗、感蓟马苜蓿叶片上、下表皮细胞纵向隆起,横向收缩。栅栏组织细胞呈圆柱状,在受害后收缩,液泡变小,单位面积细胞数增加。海绵组织细胞形态不规则,排列疏松,细胞间隙大,受害后变化不明显,且I-1健康叶与受害叶的胞间隙均大于R-1。
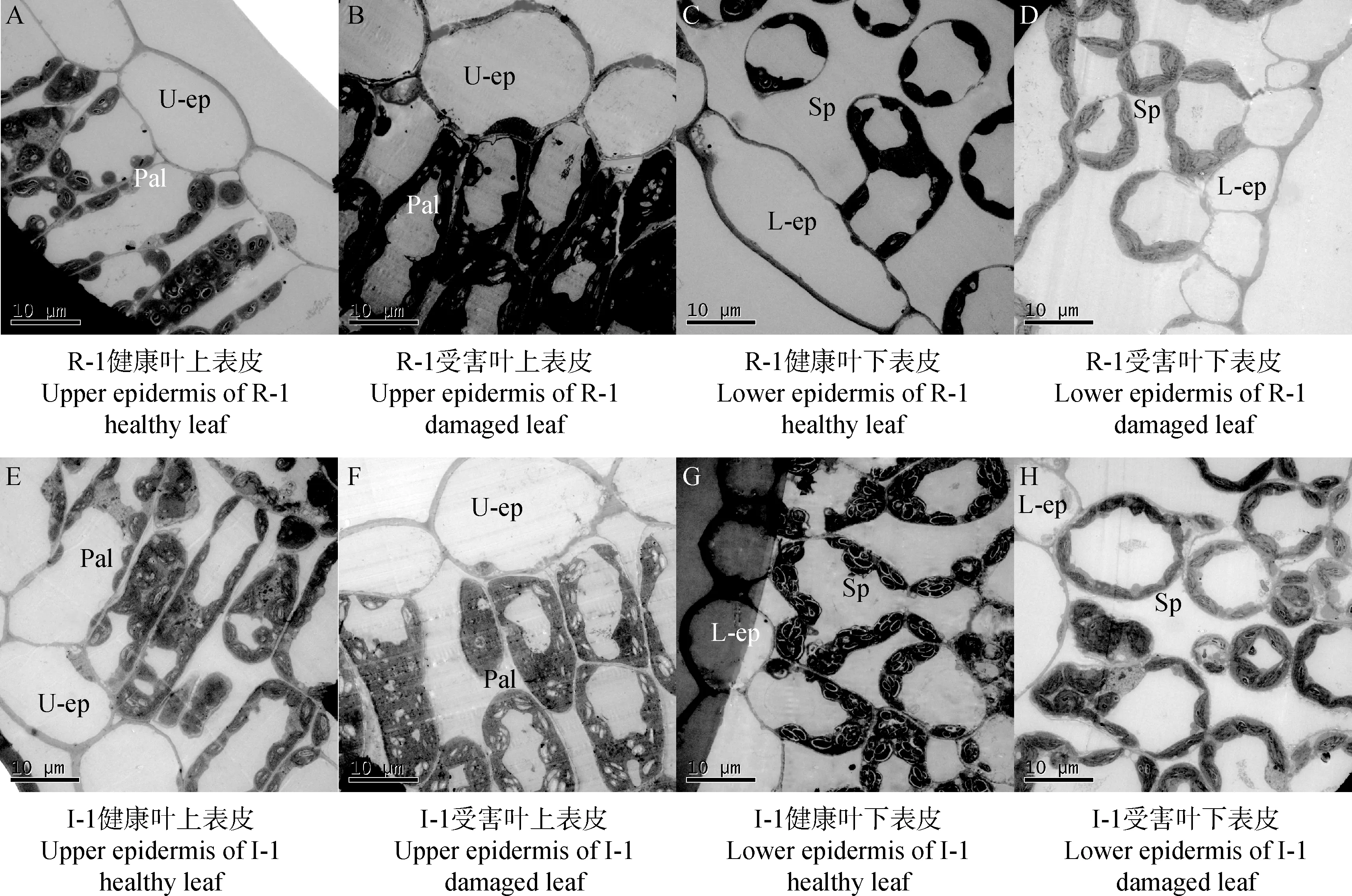
图2 R-1和I-1苜蓿健康和受害叶片表皮细胞、栅栏组织、海绵组织的变化(2500X)
2.3 抗、感苜蓿健康与受害株茎秆表皮细胞和气孔的变化
由图3可知,抗、感蓟马苜蓿茎秆平面处表皮细胞形态为不规则长方体形、排列紧密,茎秆棱角处表皮细胞形态为规则的长方形,排列更为紧密。由图3和表2可知,受蓟马为害后的R-1和I-1茎秆平面处表皮细胞体积变小,且R-1较I-1变化显著,R-1和I-1单位面积细胞数目显著增加(P<0.05),增加倍数分别为1.60和1.10。受蓟马为害后R-1和I-1茎秆棱角处表皮细胞短轴变窄,长轴变长,排列更加紧密。

表2 蓟马为害后抗、感蓟马苜蓿茎平面表皮细胞和气孔数目的变化
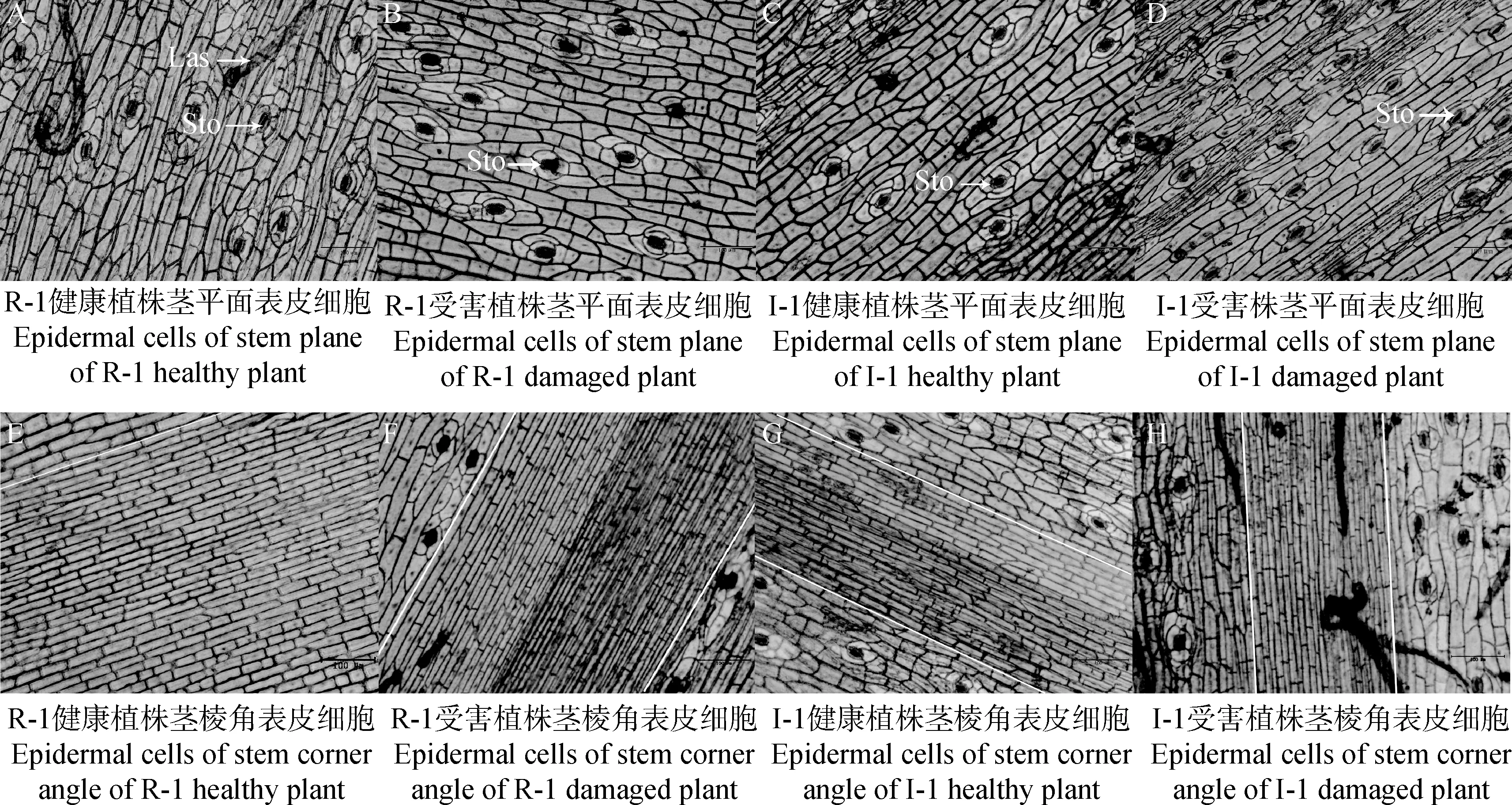
图3 健康与受害R-1和I-1苜蓿茎平面和棱角表皮细胞(10X)
2.4 抗、感苜蓿健康与受害株茎厚角组织与维管组织的变化
如图4所示,苜蓿茎的结构由表皮细胞、皮层和中柱构成,中柱中的维管束大小不一,环状排列。受蓟马为害后,抗、感蓟马苜蓿茎秆平面处的维管束间距和直径变小、维管束细胞减少;茎秆棱角处的维管束直径变小,维管束细胞减少,厚角组织的宽度也下降,且I-1比R-1更窄。

图4 健康与受害R-1和I-1苜蓿茎维管组织和厚角组织比较(10X)
2.5 抗、感苜蓿健康与受害叶肉栅栏组织超微结构的变化
健康苜蓿叶肉栅栏组织细胞叶绿体呈梭形,叶绿体膜完整,表面附着淀粉粒和少量嗜锇体。R-1健康叶片淀粉粒数量少、体积大,受害后体积变小且数量增加,嗜锇体增多。健康I-1叶片淀粉粒数量较R-1多、体积较R-1小,受害后淀粉粒体积变化不明显、数量减少,伴有嗜锇体增多。受害后I-1的嗜锇体多于R-1,抗、感蓟马苜蓿叶肉叶绿体基粒显著增加,I-1的叶绿体基粒片层和基质片层排列松散且片层间隙大(图5)。

图5 健康与受害R-1和I-1苜蓿叶片栅栏组织叶绿体电镜图片(12 000X)
3 讨论
在植物进化过程中,叶片对环境变化较为敏感,其解剖结构及微观特征最能体现环境因子的影响及植物对环境的响应[13-14]。表皮是植物抵御外界不良环境的保护组织,叶片表皮能减轻胁迫对叶片内部组织的损害[15-16]。黄亚辉等人[17]发现,茶树叶片栅栏组织和海绵组织厚度与假眼小绿叶蝉(EmpoascavitisGothe)的虫口密度极显著负相关。唐楠等[14]对青藏高原全缘叶绿绒蒿(Meconopsisintegrifolia(Maxim) Franch.)叶表皮特征的研究结果发现,随着海拔升高叶片表皮细胞形态和垂周壁式样均发生变化。陈清西等[18]通过对不同品种食用蕉(Musaacuminate×M.balbisiana)叶表皮结构的研究发现,食用蕉叶表皮细胞所具有的波浪状垂周壁,是其抗旱抗寒能力较强的表现。紫花苜蓿叶片受蓟马锉吸为害后,表皮细胞形态由不规则形变为多边形,垂周壁式样由波形变为平直,说明苜蓿叶片表皮细胞结构对蓟马的为害产生了积极响应。
周强等[19]和常金华等[20]研究认为,叶片表皮细胞和气孔密度小,以及细胞排列整齐不利于害虫的取食为害。在本试验中,抗蓟马苜蓿R-1健康叶片上下表皮单位面积细胞数目和气孔数目明显少于I-1,而在受害后,R-1叶片上、下表皮单位面积细胞数目和气孔数目增加倍数均高于I-1,且垂周壁式样由波形向平直变化,细胞排列趋向整齐,说明苜蓿表皮结构的这种变化是其对蓟马为害的抗性反应,且抗蓟马苜蓿无性系的变化更明显。
许多学者对植物显微和超显微结构研究发现,受到不利因素胁迫时,植物叶片显微结构和超微结构会受到不同程度的影响,从而影响植物的光合作用。Ivanova等[21]通过对10种不同功能类型野生植物的研究表明,所有植物类型遮荫处理后叶肉结构发生重排。陈斌等[22]对新娘草(Gibasispellucida)进行不同程度的遮光处理发现,随着光强的降低叶片栅栏组织和海绵组织的厚度显著升高。本研究发现,在受蓟马为害后,紫花苜蓿叶片栅栏组织细胞液泡缩小,细胞收缩呈椭圆形,相应地单位面积细胞数增加,而椭圆形的栅栏细胞可提高近轴面和侧面叶绿体数量,进而最大限度地进行光合作用[23]。马琳[24]的研究报道,受蓟马为害的苜蓿叶片颜色加深,叶绿素含量增加。寇江涛[25]研究表明,适当的蓟马为害可以提高苜蓿的净光合速率,使苜蓿产生光合补偿效应。说明,紫花苜蓿栅栏组织在结构上对蓟马为害产生积极响应,提高了苜蓿叶片对光能的吸收效率,进而产生光合补偿效应。
叶绿体是植物进行光合作用的主要场所,不良的生长环境会影响细胞中叶绿体的分布以及形态结构,造成基粒和片层结构数量变化、嗜锇体颗粒增加等现象[25-28]。盐胁迫时,叶绿体的响应最敏感,叶绿体超微结构发生明显变化,被膜破损甚至消失、类囊体膨大、基粒排列无序,基粒和基质片层模糊不清[29]。张洁等[30]和张桂茹等[3]研究证实了植物体的形态结构与生理功能是相统一的,单位面积细胞叶绿体数、基粒数和片层数越多,捕获光能的作用越强,光合作用越强。本试验中,受害后抗、感蓟马苜蓿叶绿体基粒均增加,R-1叶绿体基粒数和片层增加多于I-1,I-1的叶绿体基粒片层和基质片层排列松散且片层间隙大;受害后苜蓿叶肉叶绿体中的淀粉粒体积或数量减少。说明蓟马为害后,苜蓿叶片通过叶绿体基粒片层增加,提高光合效率,并将更多的淀粉转化成可溶性糖,用于植株自身的补偿生长,弥补蓟马取食为害造成的损失。
嗜锇体颗粒是类囊体降解膜脂聚集的结果,可作为植物受损程度的评价指标[31]。研究表明,高温和高光强条件下,植物叶片叶肉细胞受到损害,叶绿体上出现大量的嗜锇体颗粒[32-33]。本试验发现,取食为害后抗、感蓟马苜蓿叶肉细胞嗜锇体增多。本试验所用的叶片为受害2级苜蓿叶片,嗜锇体多少与苜蓿叶片受害程度大小之间的关系需要进一步探索。
茎是植物地上的重要部分,除了具有光合和支持作用外,最主要的功能是运输和贮存营养,保证植物正常的生长发育需求[34]。马琳等[11]研究报道,蓟马为害后苜蓿株高降低、分枝增多、节间变短、茎秆变细;受害苜蓿的同化产物更多地分配到茎秆中,用于新的侧枝和新叶生长[8],使植株产生超补偿生长,而主茎变细、节间变短可能是因为侧枝增多所致。在本试验中,苜蓿受害后茎秆表皮细胞体积变小、横切面维管束间距和直径降低,从茎秆组织显微结构的角度解释了受害苜蓿节间变短和茎秆变细的现象。植株茎秆厚角组织具有加强植株茎秆支持作用的功能[35],受害后抗、感蓟马苜蓿茎秆棱角处厚角组织宽度均有下降,厚角组织宽度的下降使苜蓿植株茎秆的支持作用变弱,而抗蓟马苜蓿厚角组织宽度下降程度较感蓟马苜蓿低,说明抗蓟马苜蓿茎秆的支持能力强于感蓟马苜蓿。
4 结论
苜蓿受蓟马为害后,叶片表皮细胞垂周壁式样由波形结构向平直变化,细胞排列趋向更加紧密和规则,单位面积细胞和气孔数目增加;受害苜蓿叶片的栅栏组织排列也更为紧密,淀粉粒减少,栅栏组织细胞叶绿体基粒与片层增加,提高了苜蓿叶片有效光合面积和对光能的吸收效率,使得苜蓿在蓟马危害后产生光合补偿,增强了苜蓿对蓟马的耐害性。R-1叶片和茎秆显微及叶片的超微结构对蓟马为害比I-1更为敏感。
猜你喜欢
江西农业大学学报(2022年3期)2022-07-07
今日农业(2021年11期)2021-11-27
少儿科技(2019年9期)2019-09-10
农业机械学报(2019年4期)2019-04-29
畜牧与饲料科学(2018年5期)2018-06-13
现代园艺(2017年21期)2018-01-03
广西林业科学(2016年1期)2016-03-20
新疆农垦科技(2014年9期)2014-02-28
土壤与作物(2013年3期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
