
饲喂PRL抑制剂对燕山绒山羊毛囊发育的影响
2021-12-31 01:39:04张乐超段春辉刘月琴张英杰
畜牧兽医学报 2021年12期
张乐超,段春辉,刘月琴,张英杰
(河北农业大学动物科技学院,保定 071000)
山羊绒是由次级毛囊(secondary hair follicle,SF)发育而来,受光照周期的调节呈季节性生长[1]。毛囊的生长周期分为生长期(5~12月)、退行期(1~2月)和休止期(3~4月),毛囊周期性再生过程是由毛囊细胞与各类信号分子交互所致[2-6],其生长机制非常复杂。皮肤是垂体外催乳素(PRL)的重要合成和靶向器官,PRL及其受体(PRLR)表达于表皮角质形成细胞、皮肤附属器、成纤维细胞、血管内皮细胞及免疫细胞中,参与调节细胞增殖与分化、毛囊周期、免疫应答等生理过程[7-9]。Foitzik等[10]研究发现,人毛囊中有丰富的PRL和PRLR,PRL可通过自分泌和旁分泌的方式调控毛囊周期性变化。研究表明,高浓度的PRL对绵羊休止期的毛囊是一种活动信号,促使处于休止期的毛囊进入毛发生长阶段,但对处于活动状态的毛囊有抑制作用[11-12]。小鼠敲除PRLR会打乱毛发周期,使毛发脱落时间提前,缩短毛囊休止期[13]。这些研究说明,PRL与毛囊周期性有关,可通过影响毛囊的生长发育调控动物毛发的生长与脱落,但关于PRL调控毛囊周期性的分子机制尚不清楚。
本研究在燕山绒山羊次级毛囊生长期(9月份)通过饲喂PRL抑制剂研究PRL对山羊绒生长及毛囊发育的影响,通过分析相关激素水平及受体表达、毛囊性状,探索PRL影响毛囊发育的机制,为揭示次级毛囊再生机制奠定理论基础。
1 材料与方法
1.1 试验地点及时间
本试验于2019年9-12月在秦皇岛青龙利红绒山羊养殖场进行。
1.2 试验设计
2019年9月,随机选取3月龄体重相近、健康良好的燕山绒山羊公羊20只,试验开始前统一用伊维菌素驱虫。试验羊随机分为2组(n=10):对照组和试验组,平均体重为(23.01±4.82)kg,试验期间试验组公羊饲喂PRL抑制剂(溴隐亭,0.06 mg·kg-1),溴隐亭溶于水中,饲喂前均匀喷洒到饲料中,对照组喷洒等量的水,试验期90 d。饲喂量的选择依据Dicks等[14]的结果(约0.07 mg·kg-1)和溴隐亭说明书(0.05~0.07 mg·kg-1)。
1.3 试验羊的饲养管理
试验期间试验羊的日粮和饲养管理与羊场其他羊只相同,全舍饲饲养。日粮由蛋白浓缩料、玉米、玉米秸秆组成,组成比例为13.5%、37.8%、48.7%。自由采食和饮水。
1.4 样品采集
饲喂PRL抑制剂3个月后,上午07:30(空腹)用5 mL真空促凝管采集两组试验羊的血液,3 000 r·min-1离心10 min后取血清样品-20 ℃保存备用。贴皮采集绒毛样品,放封口袋保存。并用一次性医用采样器(直径1 cm)采集皮肤组织2块,一块放入10%福尔马林固定液保存,另一块放入液氮保存备用。
1.5 检测方法
1.5.1 羊绒长度和细度检测 用直尺测量羊绒伸直长度,每个样品检测50根[15]。纤维细度采用全自动纤维细度分析仪BEION F10(上海北昂科学仪器有限公司)测量,每个样品检测根数>2 000根,平均3 000根左右。
1.5.2 皮肤组织染色 用HE和Sacpics方法对横切片进行染色[16]。每个皮肤样品3个重复,每例切片计数10个视野。统计初级毛囊密度、次级毛囊密度、次级毛囊与初级毛囊比值(S/P)、活性初级毛囊占比和活性次级毛囊占比。
1.5.3 激素浓度测定 将促凝管中的新鲜血液样品3 000 r·min-1离心10 min,吸取上层血清,用放射免疫试剂盒(南京建成生物工程研究所,中国,南京)测定PRL(0.5~200 ng·mL-1,CV<10%)、IGF-1(3~900 mg·L-1,CV<10%)、MT(10~3 000 ng·L-1,CV<10%)、GH(0.05~20 ng·mL-1,CV<10%)、COR(1~400 ng·mL-1,CV<10%)、T3(0.2~60 ng·mL-1,CV<10%)和T4(5~1 500 ng·mL-1,CV<10%)激素浓度。
1.5.4 皮肤总RNA提取 根据TransZol Up(Invitrogen,美国)提取总RNA,琼脂糖凝胶电泳检测,NanoDrop 2000测定RNA质量和浓度,全式金反转录试剂盒(全式金,北京,中国)进行反转录。凝胶电泳图见图1,mRNA出现明显的3条带,可用于后续试验分析。
图1 RNA凝胶电泳图Fig.1 RNA gel electrophoresis
1.5.5 RT-qPCR检测皮肤组织PRL、MTNR1a和RORα表达量 基因引物由华大公司合成,引物信息见表1。使用SYBR Green(宝生物,北京,中国)进行qPCR,利用Step One Plus仪器(Applied Biosystems,Carlsbad,CA)进行荧光检测。每孔设置3个 重复。qPCR反应体系:正、反引物各0.4 μL,Taq 酶10 μL,模板1 μL,水8.2 μL,共20 μL。qPCR程序:95 ℃ 2 min;95 ℃ 5 s、60 ℃ 30 s,40个循环;65~95 ℃,每个循环提高0.5 ℃,制作qPCR熔解曲线。

表1 试验引物信息Table 1 Primer information in this study
1.6 数据分析
采用SSCP软件中的student’sT检验分析羊绒长度、细度、毛囊数量、相关激素和基因的相对表达量(2-ΔΔCT),并统计显著性(P<0.05)。数据以“平均值±标准差”表示。
2 结 果
2.1 饲喂PRL抑制剂对山羊绒生长和绒毛品质的影响
由表2可知,饲喂PRL抑制剂3个月后对山羊绒伸直长度和细度无显著影响(P>0.05)。
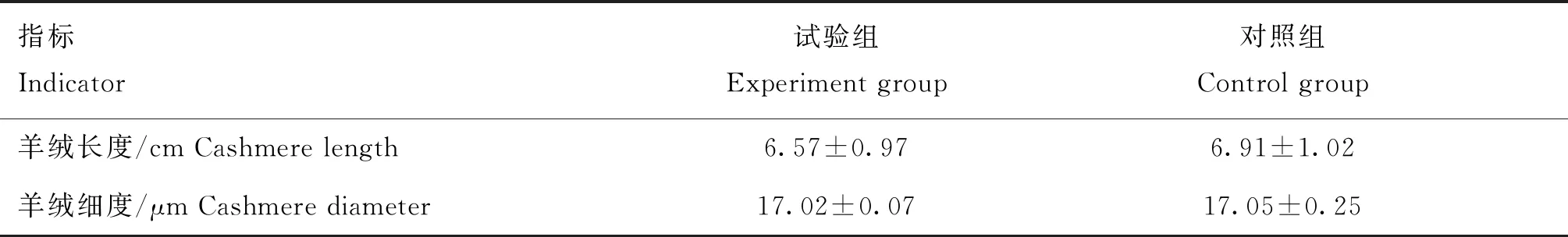
表2 饲喂PRL抑制剂对山羊绒生长和绒毛品质的影响Table 2 Effects of PRL inhibitor on growth and quality of cashmere
2.2 饲喂PRL抑制剂对绒山羊皮肤毛囊性状的影响
由图2可知,饲喂PRL抑制剂3个月显著提高了初级毛囊密度(图3A)、次级毛囊密度(图3B)、S/P值(图3C),并显著提高了活性初级毛囊占比(图3D)和活性次级毛囊占比(图3E)(P<0.05)。
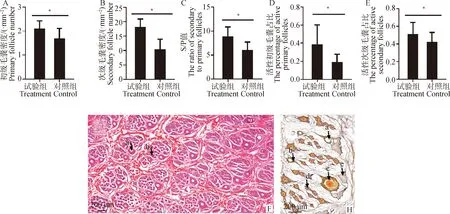
F.皮肤组织HE染色,bar =200 μm;H.皮肤组织Sacpics染色,bar =100 μm。a.初级毛囊;b.次级毛囊;c.活性初级毛囊;d.活性次级毛囊;e.皮脂腺。*.P<0.05,下同F.HE staining of skin tissue,bar =200 μm;H.Sacpics staining of skin tissue,bar =100 μm.a.Primary hair follicles;b.Secondary hair follicles;c.Active primary hair follicles;d.Active secondary hair follicles;e.Sebaceous gland.*.P<0.05,the same as below图2 饲喂PRL抑制剂对皮肤毛囊性状的影响Fig.2 Effects of PRL inhibitor on skin hair follicles
2.3 饲喂PRL抑制剂对绒山羊血清中PRL、MT、IGF-1、GH、COR、T3和T4的影响
饲喂PRL抑制剂3个月后对血清PRL、MT、IGF-1、GH、COR、T3和T4浓度无显著影响(P>0.05,图3)。
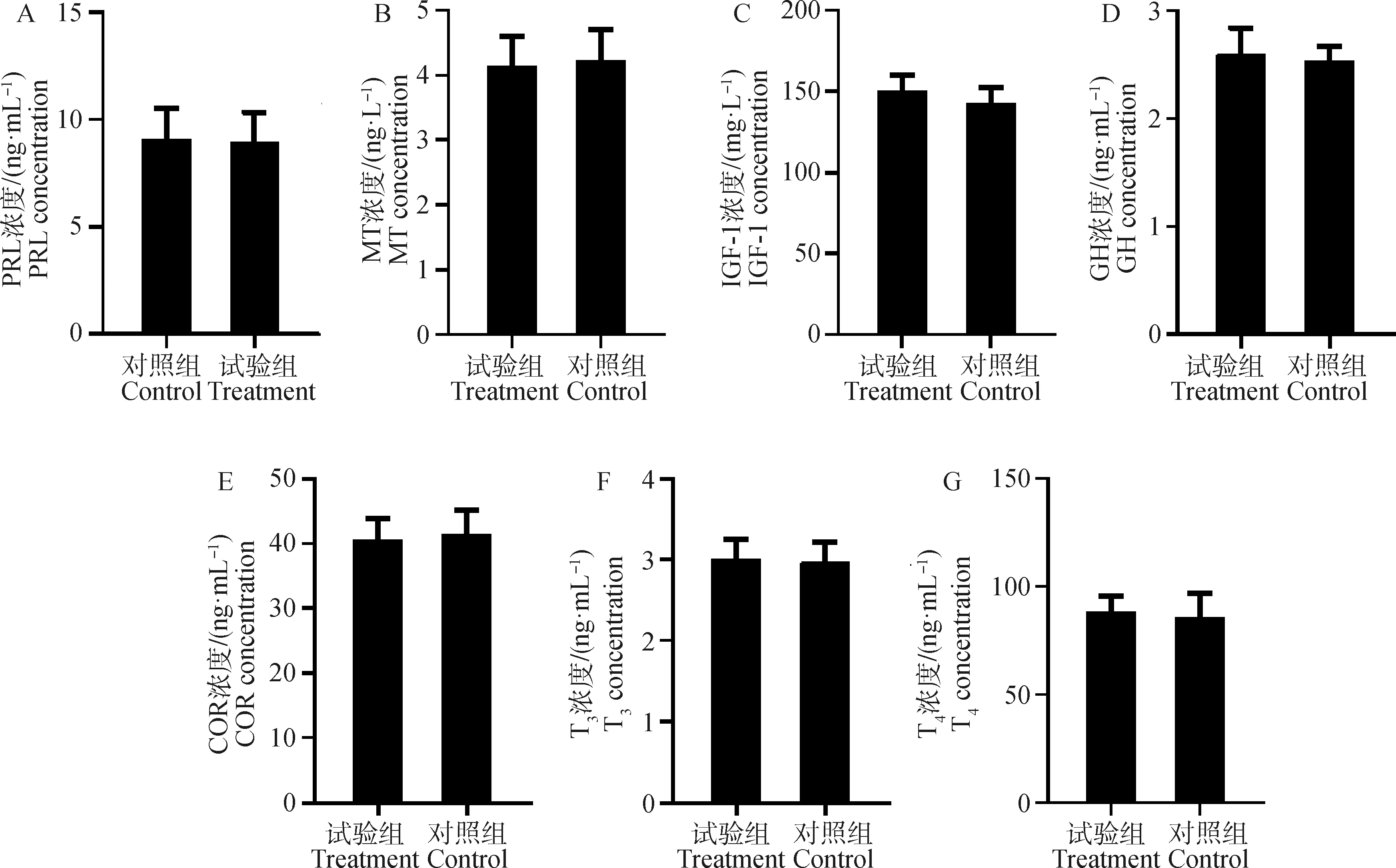
图3 饲喂PRL抑制剂对绒山羊血清中PRL、MT、IGF-1、GH、COR、T3和T4的影响Fig.3 Effects of PRL inhibitor on serum PRL,MT,IGF-1,GH,COR,T3 and T4 concentrations
2.4 饲喂PRL抑制剂对PRL、MTNR1a和RORα mRNA表达的影响
由图4可知,饲喂PRL抑制剂显著降低了绒山羊皮肤组织PRL(图4A)和MTNR1a(图4B)mRNA 表达水平(P<0.05),对皮肤组织RORα(图4C)mRNA表达量无显著影响(P>0.05)。

图4 饲喂PRL抑制剂对PRL、MTNR1a和RORα表达的影响Fig.4 Effects of PRL inhibitor on PRL,MTNR1a and RORα mRNA levels
3 讨 论
3.1 PRL对绒山羊毛囊发育及山羊绒生长的影响
Foitzik等[17]研究发现,PRL在毛囊生长期能抑制毛囊的生长,在毛囊由生长期向退行期转变的过程中,PRL的免疫活性上调,在毛囊生长末期和休止期强烈表达。另有研究发现,敲除PRLR小鼠可延长毛囊休止期,并且埋植PRL能使毛囊退行期提前,缩短毛囊生长期[13]。Nixon等[11]发现,延长光照时间可使绵羊血清PRL含量升高,次级毛囊提前进入退行期,毛囊活性降到2%。以上研究说明,高浓度PRL对毛囊生长有抑制作用,可促使毛囊由生长期向退行期转化。本研究结果发现,抑制PRL 3个月,处于毛囊生长期的羊绒长度和细度并未发生明显的变化,但试验组绒山羊皮肤中初级毛囊数量、次级毛囊数量、S/P值、初级毛囊活性占比和次级毛囊活性占比显著高于对照组,说明抑制PRL分泌后促进了生长期毛囊的发育,PRL可以调控毛囊的活化及周期性。
3.2 PRL对血清PRL、MT、IGF-1、GH、COR、T3、T4及皮肤组织MTNR1a和RORα表达的影响
Dicks等[14]发现,苏格兰绒山羊从1月开始(活化期)每2周在皮下注射35 mg溴隐亭,PRL在第1周 的第4天被显著抑制,而在第二次注射后的第11天,PRL与对照组无显著差异,持续到试验结束时的第17周。溴隐亭没有长期起到抑制PRL分泌的作用,这一现象也在赤鹿和苏格兰黑面母羊被发现[18-19]。另有研究证实,如果PRL释放被多巴胺或其激动剂抑制,PRL会在细胞分解而不分泌[20]。Blask和Orstead[21]利用多巴胺体外培养仓鼠垂体5 h,发现多巴胺可以降低培养基PRL含量,但不影响垂体3H-PRL(亮氨酸标记PRL)合成和免疫活性PRL含量。相反,Steger等[22]发现,仓鼠在接受长时间光照处理后,多巴胺并未表现出抑制PRL分泌的作用。这说明多巴胺和其激动剂可能在短期调节PRL分泌中发挥重要作用,另一个可能的解释是长期的溴隐亭处理下调了多巴胺受体的表达,多巴胺抑制作用减弱,使得PRL分泌上升[14]。Curlewis等[18]从2~11月对赤鹿注射溴隐亭,发现2~4月血清PRL含量虽然低于对照组,但差异不显著;4月中旬到9月中旬,试验组血清PRL被显著抑制,10月两组无显著差异,11月又出现明显抑制。这些变化导致粗毛和绒毛生长时间分别增加了1和3个月。因此,长期使用溴隐亭,血清PRL浓度先是无变化,然后降低,后又恢复至对照组水平,这期间的持续时间随着物种的不同而表现出长短的差异。本研究同样也观察到,溴隐亭处理3个月后,燕山绒山羊血清PRL浓度与对照组没有明显差异。本研究发现,试验组皮肤组织PRL表达量显著低于对照组,且试验组的次级毛囊数量及活性显著高于对照组,说明短期抑制PRL分泌后降低了PRL表达量,也说明皮肤中低PRL表达可促进毛囊活化,并可能会延长毛囊生长期。
大量研究表明,提高绒山羊血液中褪黑激素(MT)浓度可促进山羊绒生长、提高山羊绒产量[23-24],且PRL与MT浓度呈负相关关系[25]。本研究发现,抑制PRL 3个月后,MT浓度没有明显变化。但显著降低了MTNR1a表达量,对RORα表达量无显著影响。MTNR1a是MT膜受体的一种亚型,其表达分布对于MT功能起到重要作用,主要通过偶联G蛋白降低细胞内第二信使cAMP信号[26]。本研究检测到试验组MTNR1a表达量显著低于对照组,说明试验组细胞内酶活性要高于对照组,表明PRL可能通过降低绒山羊皮肤中MTNR1a的表达提高细胞内酶活性进而促进毛囊发育[27-29]。RORα是MT核受体的一种亚型,研究表明,RORα表达在毛囊生长期晚期较低,退行期晚期上调,休止期又下降[30]。埋植MT可以提高休止期RORα表达量,并不影响生长期和退行期的表达量[31-33]。本研究并未检测到RORα表达量发生显著变化,说明在次级毛囊生长期RORα可能不是影响次级毛囊生长发育的重要调节因子。
类胰岛素生长因子1(IGF-1)是一种多功能细胞调控因子,通过在毛囊发育过程中调节细胞增殖和迁移促进毛发生长[34-35]。绵羊皮肤注射IGF-1可增加角蛋白产生[36],缺失IGF-1受体的转基因小鼠皮肤发育不全,缺少毛囊结构[37]。另有研究发现,IGF-1转基因绵羊,羊毛纤维中副皮质细胞增加,导致直径变大[38]。且缺乏IGF-1患者的头发大部分缺乏根鞘,头发直径更细发质更脆[39]。在体外培养人皮肤毛囊的研究中发现,10和100 ng·mL-1的IGF-1 可增加毛囊长度,在有10 μg·mL-1胰岛素存在的情况下,IGF-1(0.01~100 ng·mL-1)对毛囊生长没有明显影响[40]。以上研究说明,在不同物种中IGF-1对维持皮肤功能起重要作用。本研究发现,在毛囊生长期抑制PRL分泌对IGF-1浓度无显著影响,说明在山羊绒快速生长期PRL可能不通过IGF-1对毛囊发育起作用。
皮质醇(COR)由肾上腺皮质合成和分泌,可调节免疫和应激反应[41]。Ågren等[42]发现,低水平COR可减缓皮肤结构(透明质酸和蛋白多糖)的分解,高水平COR可加快这些皮肤结构的分解。Talbott和Kraemer[43]同样发现,高水平COR可以对皮肤结构产生副作用。通过促肾上腺皮质激素释放激素(CRH)促进COR合成,可抑制体外培养人毛囊毛干产生,诱导毛囊退化,抑制角质细胞增殖[44]。本研究发现,在毛囊生长期抑制PRL分泌3个 月后,对COR浓度无显著影响,说明长期饲喂PRL抑制剂没有引起绒山羊的应激反应,且对绒山羊皮肤无损伤。而COR是否在次级毛囊发育过程中起作用还有待进一步研究。
生长激素(GH)可调节机体的生长、免疫、代谢和生殖等生理功能[45]。研究发现,GH可在真皮发挥作用,主要刺激胶原蛋白的合成,Sprague-Dawley小鼠在伽马射线照射后,皮下注射GH可保护毛囊不萎缩,真皮和表皮结构完整[46]。Celi等[47]检测绒山羊血浆GH浓度,发现GH浓度和季节无关,但高GH浓度的公羊脱绒分数要低于低GH浓度的母羊。Alam等[48]发现,100~200 ng·mL-1的重组GH(rGH)可使人毛囊提早进入退行期,显著提高细胞内IGF-1的免疫活性,且GH可通过自分泌和旁分泌的方式在靶组织中影响IGF-1发挥生理功能[49]。本研究发现,抑制PRL分泌对GH和IGF-1均无显著影响,说明在毛囊生长期抑制PRL分泌可能不会通过GH对毛囊发育起作用。
甲状腺激素由甲状腺分泌,可调节动物机体的生长发育、组织分化和物质代谢等。甲状腺激素主要有T3和T4,二者分泌呈季节性变化[47]。Vidali等[50]用100 pmol·L-1T3和100 nmol·L-1T4分别处理人的毛囊,发现二者减少了活性氧的形成,增加了活性氧清除剂(过氧化氢酶和超氧化物歧化酶2)的转录,刺激毛囊角质细胞产生ATP。Villar等[51]发现,山羊皮肤可发生甲状腺激素代谢,且甲状腺激素代谢与脱碘酶(MD)活性有直接关系,推测皮肤MD活性可能与羊绒生长有关;其在绒山羊皮肤组织检测到了MDII和MDIII,MDII可使T4转化为T3,MDIII可使T3转化为T2、T4转化为rT3,二者活性可能对脱绒模式有影响[51]。Merchant和Riach[52]发现,高水平的甲状腺激素可以延迟脱绒时间和毛囊的活化。Celi等[47]检测1年甲状腺激素(T3和T4)水平,发现T4与PRL有正相关关系,T3与脱绒有正相关关系,并且在2月份达到最高水平,11月份达到最低水平。本研究发现,试验组和对照组T3和T4并没有显著差异,说明在山羊绒生长期抑制PRL分泌对甲状腺激素分泌无显著影响。
4 结 论
本研究结果显示,PRL可调控绒山羊毛囊的发育,在绒山羊毛囊生长期抑制PRL分泌可促进毛囊发育,提高毛囊数量和活性毛囊比例,且皮肤中PRL和MTNR1a基因在PRL调控毛囊发育中起重要作用。
猜你喜欢
毛纺科技(2023年12期)2024-01-11 01:46:10
军事文摘(2023年2期)2023-02-17 09:20:24
今日农业(2022年14期)2022-09-15 01:43:28
纺织标准与质量(2022年3期)2022-08-10 09:11:14
感染、炎症、修复(2021年1期)2021-07-28 06:18:08
东华大学学报(自然科学版)(2018年1期)2018-06-29 03:34:50
新农业(2016年23期)2016-08-16 03:38:12
新农业(2016年22期)2016-08-16 03:34:42
保健与生活(2016年1期)2016-04-12 18:29:44
中国纤检(2015年15期)2015-11-13 18:21:58
