
三峡水库坝下江心洲植物群落及其性状对不同水淹强度的响应
2021-12-24 09:55孙小祥易雪梅黄远洋冉义国罗高行马茂华吴胜军
生态学报 2021年23期
孙小祥,易雪梅,黄远洋,冉义国 ,罗高行,马茂华,吴胜军
1 中国科学院重庆绿色智能技术研究院,重庆 400417 2 中国科学院水库水环境重点实验室,重庆 400417 3 中国科学院大学,北京 100049 4 重庆交通大学,重庆 400074
三峡大坝建成运行后,发挥了对坝下游长江流域的防洪和供水安全保障的重要作用。与此同时,坝下游水沙情势发生了显著变化,宜昌站年输沙量较建坝前多年平均输沙量减少85%,下泄水流的挟沙量长期处于未饱和状态,形成清水下泄的水沙情势[1]。坝下游河床和洲滩等河流地貌通过长期的自动调整以适应清水下泄的水沙情势[2- 3],使河流能以最小的能量输送泥沙荷载[4]。其直接后果是严重影响下游江心洲发育,引起洲体面积减小和整体蚀退,对河槽稳定、沙洲湿地和农业生产造成严重影响[5]。
坝下游洲滩植被作为洲滩生态系统的重要组成部分,其空间分布和稳定性是维持洲滩生态系统功能的关键[6- 7]。在洲滩适应清水下泄水沙情势的过程中,不仅洲滩的冲淤过程影响植被分布[8],植被分布也同时对洲滩的冲淤过程具有重要影响[9- 10],进而对洲滩的形态调整产生反馈作用。受大坝运行影响,坝下生态系统成为响应水文形势变化的敏感区域[11],特别是周期性的水文调控对坝下生物区系产生了巨大影响[12- 13]。本研究选取对水沙情势变化响应十分敏感的太平口心滩作为研究地点,结合洲滩水淹强度,通过研究江心洲植物群落结构、空间分布及性状特征,探讨坝下游沙洲植物群落构建过程,以期为坝下游沙洲发育机制研究、洲滩生态工程及洲滩农业等提供研究基础。
1 研究地区与研究方法
1.1 研究区概况
研究区域太平口心滩(30°30′N,112°13′E),位于湖北省荆州市荆江河段中的上荆江河段。荆江河段是受大坝调蓄水影响最为显著的河段之一[14- 15],同时是三峡坝下游第一个沙质河床河段,而太平口心滩又是该河段上游的第一个江心洲,因此太平口心滩对水沙情势变化的响应十分敏感。该区域距离三峡大坝下游约180 km,地处亚热带季风气候区域,气温季节变化显著,四季分明。全年平均温度为14—22℃;降水受季风影响,季节性强并且集中,年际变化显著,年均降水量1098;无霜期年均值为268 d,占全年总日数的73%。太平口心滩最早在20世纪70年代由腊林洲边滩头部切割发育而来[16],经历了近50年的发育。太平口心滩因为从形成到现在发育时间较短,且离河岸较远,受人为干扰小。因此太平口心滩是一个具有植被原生演替特征的江心洲。
1.2 样地设置及调查
结合心滩不同时期的出露情况,在地理信息系统ArcGIS中WGS84+UTM48N坐标系统下对样带和固定样方进行事先设定。根据心滩面积,设置50 m×50 m控制网格,在每个50 m×50 m控制网格中,选取规则分布的5 m×5 m样方进行调查,从而形成以25 m为间隔的梯度样方,对心滩进行系统性调查及采样。植被调查在2019年12月完成。野外观测时,采用RTK(Real-time kinematic) 对样方进行放样,并记录高程信息。植被调查记录到有植物生长的样方共计55个。记录样方内出现的植物物种名称,并测量其高度、盖度以及数量。同时记录样方地形、土壤类型及地表覆被等情况。通过《中国植物志》(http://www.iplant.cn/frps)查询记录到的植物物种的生活型。野外调查时不能直接识别的,记录到所属的科。样方布设与植被生长情况如图1。
1.3 水淹梯度的划分
根据RTK记录的样方高程信息以及太平口心滩2019年的水位数据,通过调查日的水位找到在水位上涨前具有相同稳定水位的日期,作为一个水淹周期的起始(图2)。通过一个水淹周期的水位和样方的高程信息,得到每个样方在一个水淹周期内被淹没的天数。通过水淹天数将样方划分为5个水淹时间梯度,五个梯度的水淹天数分别为:20—40 d、40—60 d、60—80 d、80—100 d、100+ d。

图2 2019年太平口心滩水位趋势图Fig.2 The water level of Taipingkou Central Bar in 2019
1.4 数据的分析处理
1.4.1重要值
选取重要值(Important value,Iv)作为衡量心滩各物种相对重要性的综合指标[17]。因本研究前期勘测发现心滩植被以草本植物占据绝大多数,只有零星川三蕊柳分布,川三蕊柳属于灌木或小乔木,因此未设置乔木样方。川三蕊柳的重要值计算也按照草本植物的计算方式得出。
重要值(Iv)=(相对盖度+相对频度+相对密度)/3
式中,相对密度 =(某种植物的个体数/全部植物的个体数)/100%
相对频度 =(某个种在统计样方中出现的次数/所有种出现的次数)/100%
相对盖度 =(某个种在统计样方中的总盖度/所有种的总盖度)/100%
1.4.2生物多样性指数
生物多样性指数包括:物种丰富度S、Shannon-Weiner指数、Pielou均匀度指数、Simpson优势度指数[18]。
物种丰富度:
S=样方内物种数
Pielou均匀度指数:E=H/ln(S)
式中,S代表出现的物种个数;pi表示第i个种的多度比例;ni表示第i个物种的个体数;n表示出现的物种个体总数。
1.4.3功能多样性指数
本研究选取植株高度、生活型、花果期、根系类型及是否进行营养繁殖等一系列可以表征植物功能性状的特征进行功能多样性指数的计算(表1)。功能多样性指数包括:功能丰富度(Functional Richness;FRic)、功能均匀度(Functional Evenness;FEve)、功能分异度(Functional Divergence;FDiv)。以及群落功能加权平均值(Community-Weighted Mean values,CWM)体现不同群落间单个功能性状的差异。功能多样性指数和CWM值都通过R 4.0.2(FD Packages)得到。需要注意的是三个功能多样性指数对于物种数小于3的样方无法计算得到。

表1 植物的功能性状
本研究所有的数据处理与可视化均使用Excel 2016、R 4.0.2以及Past3软件进行。
采用单因素方差分析(one-way ANOVA)讨论不同水淹梯度下生物多样性指数的显著性差异,并通过LSD法进行多重比较。通过非参数多元方差分析(Permutational multivariate analysis of variance,PERMANOVA)检验不同水淹梯度下植物物种组成是否有显著性差异,检验过程基于Bray-Curtis相似性矩阵,通过999次置换分析确定显著性。并采用两两比较和相似百分比(SIMPER)指数寻找对群落变化具有较高贡献率(≥1%)的物种,在二维排序空间使用NMDS展现。对功能多样性指数进行沿水淹梯度的分布统计,并通过对不同样地的功能加权平均值[19]和样方信息进行非约束排序,探讨植被不同功能性状对于淹水时间的响应关系。
2 结果与分析
2.1 植物物种组成
此次对太平口心滩样方调查发现和确认草本植被群落共有21科33属39种(表2),其中:禾本科(Poaceae)10种9属;菊科(Compositae)8种8属;豆科(Fabaceae)3种3属;苋科(Amaranthaceae)2种2属;其他科:如唇形科(Labiatae)、桔梗科(Campanulaceae)、马齿苋科(Portulacaceae)、木贼科(Equisetaceae)、茜草科(Rubiaceae)、伞形科(Umbelliferae)、莎草科(Cyperaceae)、十字花科(Brassicaceae)、景天科(Crassulaceae)、石竹科(Caryophyllaceae)、蓼科(Polygonaceae)、通泉草科(Mazaceae)、碗蕨科(Dennstaedtiaceae)、杨柳科(Salicaceae)、紫草科(Boraginaceae)、酢浆草科(Oxalidaceae)均为1属1种。禾本科植物最丰富,其次分别是菊科植物,豆科植物和木贼科植物,分别占此区域草本总科数的59.6%、12.1%、6.7%和6.4%,是该区的主要优势科。其他科优势度不明显,且单属、单种现象明显。

表2 太平口心滩植物群落的物种组成
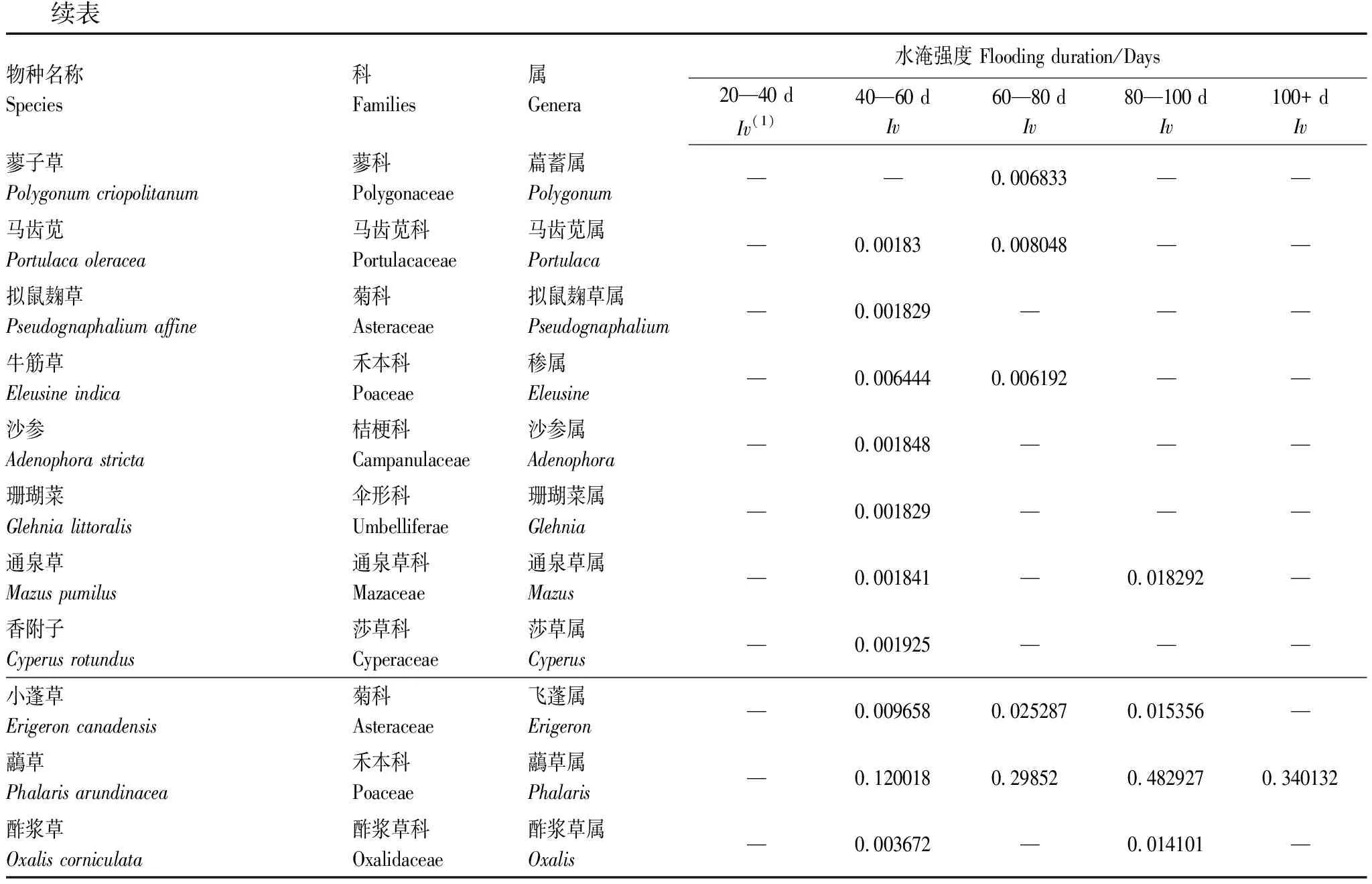
续表物种名称Species科Families属Genera水淹强度 Flooding duration/Days20—40 dIv(1)40—60 dIv60—80 dIv80—100 dIv100+ dIv蓼子草Polygonum criopolitanum蓼科Polygonaceae萹蓄属Polygonum——0.006833——马齿苋Portulaca oleracea马齿苋科 Portulacaceae马齿苋属Portulaca—0.001830.008048——拟鼠麹草Pseudognaphalium affine菊科Asteraceae拟鼠麹草属Pseudognaphalium—0.001829———牛筋草Eleusine indica禾本科Poaceae属Eleusine—0.0064440.006192——沙参Adenophora stricta桔梗科Campanulaceae沙参属Adenophora—0.001848———珊瑚菜Glehnia littoralis伞形科 Umbelliferae珊瑚菜属Glehnia—0.001829———通泉草Mazus pumilus通泉草科Mazaceae通泉草属Mazus—0.001841—0.018292—香附子Cyperus rotundus莎草科Cyperaceae莎草属Cyperus—0.001925———小蓬草Erigeron canadensis菊科Asteraceae飞蓬属Erigeron—0.0096580.0252870.015356—虉草Phalaris arundinacea禾本科Poaceae虉草属Phalaris—0.1200180.298520.4829270.340132酢浆草Oxalis corniculata酢浆草科Oxalidaceae酢浆草属Oxalis—0.003672—0.014101—
太平口心滩植物群落优势物种为虉草、牛鞭草、芦苇、狗牙根和蒌蒿,占所有物种的56.18%。水淹强度超过40 d后,随着水淹强度的增加物种数有所降低,物种数最少的是水淹强度处于100+ d的生境,仅出现了8种植物,且以虉草和狗牙根作为优势物种。物种数最多的是水淹强度为40—60 d的生境,调查发现的共39种植物中出现了35种,优势物种为牛鞭草和虉草。水淹强度为80—100 d、60—80 d和20—40 d的生境物种数分别为15、18和12种。其中水淹强度为80—100 d和60—80 d的生境优势物种为虉草和牛鞭草;20—40 d生境优势物种为牛鞭草和节节草。可以看出,虉草在整个心滩都具有优势,且随着水淹强度的减弱虉草优势度有所下降。随着水淹强度的减弱,斑茅和芦苇开始出现在水淹强度为80—100 d的生境;荻在水淹强度为60—80 d的生境开始出现;水淹强度最弱的20—40 d生境相较于40—60 d生境反而有许多物种未出现。
2.2 不同水淹强度下植物群落物种多样性
根据各样方在一个水位周期被淹没的天数和各样方记录的物种数据,分析物种多样性随着水淹强度的变化,得到心滩植物群落的物种多样性指数沿水淹强度梯度的变化。由图3可以看出,在不同的水淹梯度下,太平口心滩的植物群落物种丰富度、Shannon-Weiner指数、Simpson指数和均匀度指数均存在显著性差异。其中物种丰富度随着水淹强度的增加呈先增加后降低的趋势,并在40—60 d达到峰值,显著高于处于其他水淹强度的生境,物种丰富度在水淹强度最大的100+ d处达到最低值,显著低于其他生境。Shannon-Weiner指数和Simpson指数具有相似的趋势,随着水淹强度的增加,先缓慢下降后急剧降低。Shannon-Weiner指数和Simpson指数在水淹强度为20—40 d和40—60 d时显著高于其他生境,且在20—40 d时最高,水淹强度为100+ d时显著低于水淹强度小于100天的生境。物种均匀度趋势在水淹强度为100+ d时达到最大,在水淹强度为40—60 d时最小。

图3 太平口心滩不同水淹强度下物种多样性格局Fig.3 Species diversity of different flood duration in Taipingkou Central Bar图中小写字母表示生物多样性指数在不同水淹强度下的差异性(P<0.05)
2.3 不同水淹梯度下植物物种组成特征
PERMANOVA检验结果显示,不同水淹梯度下植物物种组成具有显著性差异,并进行两两比较(表3)。通过Simper分析得到物种对于群落间变异的解释率。使用非度量多维尺度分析在排序空间展示组间差异如图4(Stress=0.22)所示。
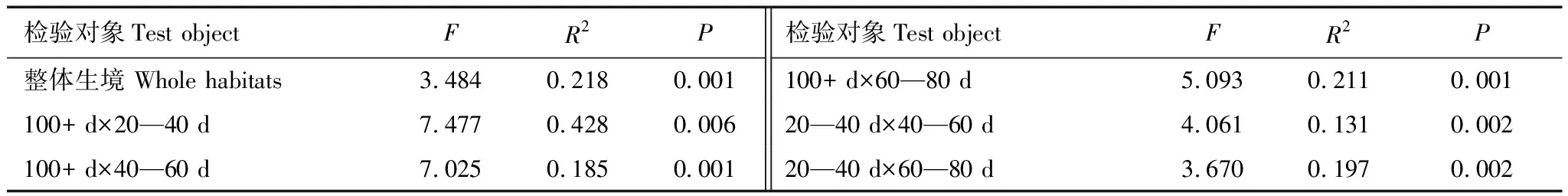
表3 PERMANOVA和两两比较显著性差异检验表
NMDS排序图显示,水淹强度为100+ d和20—40 d与其他水淹天数下的植物群落组成可能具有显著性差异。通过两两比较结果验证得到:水淹强度100+ d与20—40 d有显著性差异,且节节草、芦苇和牛鞭草解释了84.3%的群落变异;水淹强度为100+ d与40—60 d有显著性差异,牛鞭草、虉草、荻、狗牙根四种植物贡献了70.5%的群落变异;水淹强度为100+ d与60—80 d有显著性差异,虉草、芦苇、牛鞭草、狗牙根四种植物贡献了79.4%的群落变异;水淹强度为20—40 d与40—60 d有显著性差异,节节草、芦苇、牛鞭草三种植物贡献了52.1%的群落变异;水淹强度为20—40 d与60—80 d有显著性差异,节节草、芦苇、虉草三种植物贡献了57.8%的群落变异。

图4 太平口心滩植物群落非度量多维尺度分析(NMDS)排序图Fig.4 Non-metric multidimensional scaling(NMDS) of plant communities in Taipingkou central bar
2.4 植物群落功能特征对不同水淹强度的响应
不同水淹强度下的植物群落功能多样性特征如图5。水淹强度100+ d的8个植物群落,有7个物种组成都小于三种,因此无法得到功能多样性指数的值。除去水淹强度为100+ d的生境,可以看出功能丰富度指数(FRic)随着水淹强度的增加而降低,水淹强度20—40 d时Fric最高(0.21),水淹强度为60—80 d和80—100 d时Fric值达到最低(0.1)。功能均匀度指数(FEve)随着水淹强度的增加呈先下降后上升的单峰分布格局,且在水淹强度为20—40 d达到最大(0.48),水淹强度为40—60+ d时达到最低(0.26)。功能分异度指数(FDiv)随着水淹强度的增加呈缓慢下降的趋势,水淹强度为20—40 d时功能分异度指数最大(0.95),水淹强度为80—100 d时FDiv最小(0.74)。
RDA全模型检验结果表明模型成立[20],前两轴共计解释了23.48%的特征变量。解释变量与前两轴的显著性检验发现物种生活史形态与响应变量差异不显著,其他性状均具有显著性差异。结合前向选择对余下功能特征的CWM值进行排序(图5)。随着水淹强度的增加,心滩植被的花期相对位于出露期,同时植株高度更加低矮,植物根系也随着水淹的加强偏向于须根系。植被是否进行营养繁殖同水淹强度变化的响应关系较弱,可能会随着水淹强度的加强偏向于营养繁殖的方式。

图5 太平口心滩不同水淹强度下植物功能多样性格局Fig.5 Species Functional diversity of different flood duration in Taipingkou Central BarFRic:功能丰富度 functional richness;FEve:功能均匀度 functional evenness;FDiv:功能分异度 functional divergence
3 讨论
对太平口心滩的植被调查发现:心滩植物组成以草本植物为主,未见乔木,稀见川三蕊柳,有高大草本荻和斑茅分布。表明心滩植物群落整体处于演替的草本阶段,这也符合心滩的原生演替特征和洲体较短的形成发育时间特点。心滩植物群落的优势科属为禾本科植物和菊科植物,因为周期性的水淹作为一个环境过滤因子,对江心洲植物群落具有一定的选择性。菊科植物的进化程度较高,可以通过繁殖体优势和抗逆等特征适应心滩生境,禾本科植物是湿地生境的常见优势植物[21- 22]。牛鞭草、芦苇、狗牙根和蒌蒿是太平口心滩的优势物种,不同水淹强度下的植物群落优势物种不一样,水淹强度超过60 d的三个生境虉草优势度最高,低于60 d的两个生境牛鞭草优势度最高,表明在整体环境过滤的条件下,江心洲植物群落还在水淹强度梯度下具有一定程度的分化。
太平口心滩植物群落沿水淹强度的梯度特征不但体现在物种组成上,也体现在群落结构上。群落物种多样性(丰富度、Shannon指数和Simpson指数)都在100+ d达到最低值,均匀度达到最大值。这是因为在极端水淹强度下,环境过滤效应强,只有虉草和牛鞭草等适应极强水淹条件的先锋物种可以存活,导致物种多样性低[23- 24]、均匀度高。在物种组成方面,100+ d和20—40 d的生境物种组成显著区别于其他三种处于中度水淹强度的生境(40—100 d),且100+ d和20—40 d生境下的物种数都要少于中度水淹条件,这可能是因为随着水淹强度的减弱,在20—40 d生境中,物种间的竞争效应加强[25]。这符合Bertness等[25]等提出的胁迫梯度假说,即物种间的互惠关系随着胁迫强度的增加而增加。这也解释了水淹强度最弱的20—40 d生境物种数少于中度水淹条件,但是物种多样性却保持最高。后有学者对胁迫梯度假说作出补充,即在极端胁迫条件下种间互惠效应有所减弱,出现具有相似性状的物种竞争这一情况[26]。这和极端水淹生境下的100+ d群落物种均匀度高、异质性低这一现象基本一致。中度水淹强度下的生境物种种类最多,小灌木川三蕊柳和高大草本荻、斑茅也在中度水淹生境中出现。这一现象可以用中度干扰假说(The intermediate disturbance hyphothesis, IDH)解释,即生物群落在中度扰动的情况下会保持物种物种丰富度最高[27]。结合以上研究结果和理论分析,可以看出在水淹强度呈梯度变化的太平口心滩,极端水淹强度下环境过滤主导群落的构建过程,导致其生物多样性低,随着水淹强度的降低,强烈的非生物胁迫环境有所改善,植物群落受到的环境过滤效应减弱,中性构建过程和种间互惠作用加强,物种多样性增加,当水淹强度达到20—40 d时,和100+ d生境下的环境过滤效应主导相比,转换为种间竞争主导的构建过程。
太平口心滩植物群落功能多样性除去水淹强度最强的100+ d,不同水淹强度下的群落功能多样性特征(FRic和Feve)分布趋势和物种多样性趋势相同,即功能丰富度(FRic)随水淹强度的增加而单调降低,功能均匀度(FEve)随水淹强度的增加先降低后增加。功能丰富度本身和物种多样性呈正相关的关系[28],功能均匀度和功能分异度(FDiv)与物种多样性之间是相互独立的[29]。功能分异度在不同水淹强度生境的区别不大,这可能和本次研究选取的几种功能性状都和植被适应周期性水淹有关,周期性水淹作为太平口心滩整体的一个环境过滤因子,这几种功能性状在不同水淹强度生境下都有出现。
对群落功能加权平均值(CWM)的RDA的排序结果表明,随着水淹强度的增加,植被更倾向于表现出花期和心滩出露期重合度高、植株高度降低、须根系、进行营养繁殖的功能性状。植被花期和太平口心滩出露期的高度重合可以保证植物在有限的出露时间内完成自己的生活史发育。植株的高度随水淹强度的增加更加低矮可能是因为植物需要在极短的时间内完成自己的生长发育,植物将更多的资源用于生殖生长而不是营养生长。这一现象也体现在随着心滩植物花期的到来,植物的营养生长受限(高度低矮、须根),这是因为植株在胁迫条件下会将资源更多的分配给生殖生长,确保其繁衍生存,属于逆境下植物资源分配的适应策略[30]。同时须根系的植物可以更好的利用流动性资源,进行营养繁殖的植物可以迅速占领生境,使植被在心滩出露期快速建立优势。以上这些植物性状都可以让植株更好的利用洪水过程所带来的流动性资源[31- 33],表现为植物可以在周期性的出露窗口迅速完成自己的生活史周期,具有很强的可塑性,而不是对洪水过程采取抵抗性策略(更高的株高等)[34]。
值得注意的是,本研究中处于水淹强度最强的100+ d生境中的8个样方,除去一个样方内记录物种数达到7种以外,其他七个样方物种数均为1—2种,未能计算出该生境的功能多样性指数(FRic,FEve,FDiv)。但是通过比较该异常样方(物种数达到7)的年际高程信息,发现其2019年吴淞高程为35.61 m,而在2017年为39.52 m,年际高程差接近4 m。这表明心滩确实处于严重蚀退,同时该样方群落显著区别于同年相同水淹强度的其他样方,这一现象是否与其在调查年之前物种丰富度本身较高有关尚不清楚。由此延伸出来的关于周期性扰动下江心洲植物群落动态与构建过程问题需要获得更多的关注。
4 结论
在周期性水淹的胁迫干扰下,江心洲植物群落整体受环境过滤效应的主导,随着水淹时间强度的变化,不仅在物种尺度具有与之对应的梯度特征,性状组成和结构方面也具有明显的对应关系。就植物群落功能特征而言,一系列可以帮助植物快速获取流动性资源的性状更加适合水淹较强的区域,包括花果期和出露期的重合度更高、须根系、植物高度更加低矮和进行营养繁殖。对于周期性水淹梯度下的江心洲而言,主导不同水淹时间梯度下植物群落特征的生态学效应不同,江心洲植物群落物种组成及功能性状特征梯度效应明显。
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
电影评介(2022年4期)2022-08-08
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
华人时刊(2020年17期)2020-12-14
当代陕西(2020年20期)2020-11-27
汽车实用技术(2019年20期)2019-11-26
商周刊(2019年18期)2019-10-12
中国石油大学胜利学院学报(2019年2期)2019-07-08
