
不同C/N比对杉木凋落叶分解过程养分释放速率的影响
2021-12-23 07:29陈义堂
四川林业科技 2021年6期
陈义堂
福建省洋口国有林场, 福建 顺昌 353211
凋落物亦称枯落物,是生物组分产生并且归还到地面的新陈代谢产物,其发生在植物生长发育过程中[1]。它是降解者的物质和能量来源,影响陆地生态系统净生产力,在维持土壤肥力,对物质循环和能量流动有不可或缺的作用[2-3]。目前有关凋落物分解的研究主要集中于2个方面,一是关于凋落物所处的外在分解环境[4-5],二是凋落物自身的质量,即关于凋落物自身的化学性质和养分含量[6-7]。氮作为植物生长所必要的重要元素之一,同样也是构成凋落物质量的重要指标,凋落物以及所在环境中氮元素有效性的高低同样对凋落物的分解具有重要作用。Vivanco等[8]和项文化等[9]的研究结果表明施用外源氮可以促进凋落物的分解。已有研究证明,凋落物的分解速率与其自身所含的初始C/N比值存在显著的线性关系[10,11]。大量的研究结果表明,杉木凋落物在分解初期阶段普遍存在着氮富集现象,杉木新鲜凋落物中的氮养分含量较低,难以维持分解者的生长发育,从而阻碍了杉木凋落物的分解,其分解速率常与氮以及其相关的凋落物质量指标,如C/N和木质素/N比值等指标具有紧密相关性,而通过外源性施氮处理,则有利于杉木凋落物的分解[12]。因此,大多数学者提议将凋落物本身的C/N比值作为衡量其质量和预测其分解速率的重要指标[13],并推测凋落物初始养分中含有某个C/N比临界值,如果高于这一C/N比值则不利于微生物对凋落物的分解和利用,导致养分被强烈固持于凋落物中[14-15],反之则有利于凋落物的分解。
杉木(Cunninghamia lanceolata)是我国南方主要的速生针叶用材树种之一,具有生长快,材质好,病害少等优点[16]。随着社会的发展,林业集约生产水平逐渐提高,杉木速生、丰产的优点得到充分地体现,但是由于人工林连栽造成的地力衰退现象已经成为影响杉木人工林可持续经营的主要因素[17]。除了采取人为措施恢复地力外,土壤的培肥能力也是关键因素之一,凋落物的分解作为土壤养分的重要来源之一,对土壤培肥功能的研究至关重要,因此,加强杉木凋落物分解特性的研究,对于维持与提高杉木人工林地力具有重要意义。本研究以福建省南平市顺昌洋口国有林场南山管护站杉木大径材林分为试验地,研究林下植被保留和林下植被去除2种林下植被管理措施下不同C/N比的杉木凋落叶分解情况,通过外源氮添加调控杉木凋落叶的C/N比值,并置于上述2种林分中自然分解,观测其分解过程中碳、氮、磷、钾、钙、镁和锰的养分动态和释放规律以及其相关性。以期进一步论证杉木凋落物分解的内在C/N比值机理,从而为促进杉木人工林的养分循环,改善土壤养分状况,缓解杉木人工林地力下降速度、维持杉木人工林长期生产力提供科学理论依据。
1 试验林概况
试验地位于福建省南平市顺昌县洋口国有林场南山工区下林厂(117°92′~118°64′E,26°87′~27°13′N),属于中亚热带季风湿润气候,年均气温18.6 ℃,年均降水量1 600~1 900 mm,年均蒸发量为1 308~1 587 mm,相对湿度平均值为78%~82%,全年日照时数为1 668~1 972 h,无霜期254~305 d,试验林海拔252~339 m,土壤为山地红壤。
林下植物保留及林下植被去除试验林为相互毗邻的林分,且均于1985年采用杉木初级种子园的优良家系壮苗造林,造林密度为2 505 株·hm-2。1997年8月进行抚育间伐,林分保留密度为1 187 株·hm-2。间伐后,林下植物保留林分采用近自然经营的林下植被管理方式保留林下植被;经2018年6月林下植被调查,林下灌木层主要为:大叶紫珠(Callicarpa kochiana)、杜茎山(Maesa japonica)和粗叶榕(Ficus hirta)等,林下草本主要为薄盖短肠蕨(Allantodiahachijoensi)和金毛狗(Cibotium barometz)等,草本层生物量6.70 t·hm-2,林下植被总盖度92%。林下植被去除林分自2012年7月起,每年进行1次林下植被清理,林下灌木清除采用割灌机,林下草本采用镰刀手工割除。
2 研究方法
2.1 样地设置及取样
2018年11月在2种林分下分别设置3个20 m×20 m样地,对每块标准样地内的杉木进行每木调查,采用围径尺测量胸径,红外线测高测距仪测量树高,并在样地的4个角均打下水泥桩以作标记。不同林分类型样地基本情况见表1。
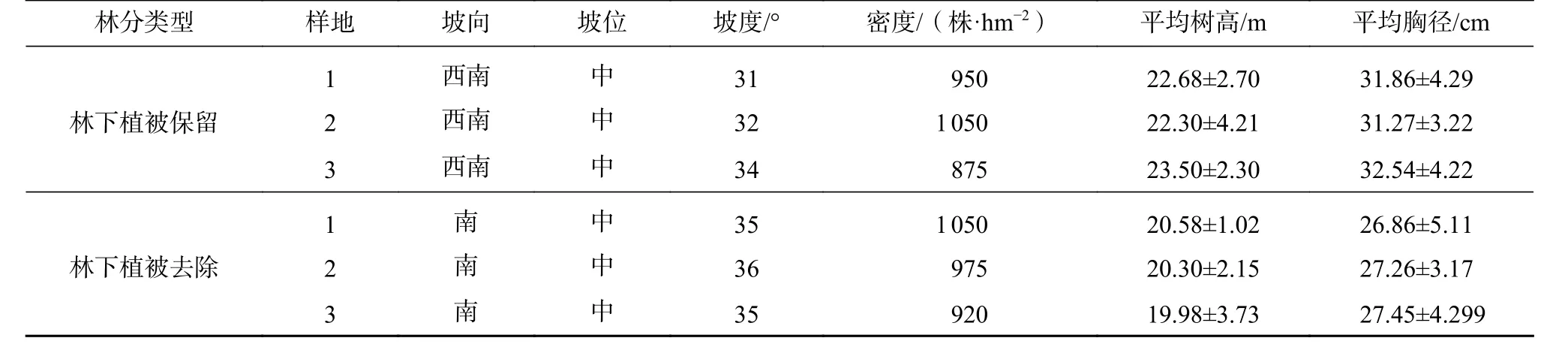
表1 试验林分样地基本情况Tab.1 General information of sample plots with different understory vegetation management measures
为保证凋落叶养分含量基本一致采用异地分解法。试验材料取自杉木中龄林尚未分解的新鲜杉木凋落枝,将杉木凋落叶从收集的杉木凋落枝剥离后,带回试验室自然风干后备用。经测定,杉木凋落叶初始全氮含量为8.10 mg·g-1,全碳含量为482.54 mg·g-1,全磷含量为0.33 mg·g-1,C/N比值为60.86,C/P比值为1 393.19,N/P比值为23.86。杉木凋落叶初始钾含量为2.89 g·kg-1,初始钙含量为4.23 g·kg-1,初始镁含量为1.33 g·kg-1,初始锰含量为1.21 g·kg-1。
2.2 凋落叶分解袋的放置、取样和预处理
试验共设计了4种C/N处理:C/N比值60.86(CK)、C/N比值40.57(N1)、C/N比值30.45(N2)、C/N比值20.25(N3),每个处理重复25次。选取NaNO3作为外源氮源,分别将质量浓度为2 g·L-1、4 g·L-1和6 g·L-1的NaNO3溶液均匀喷施于叶面,少量多次,待其风干后测定C/N比值含量,直至各处理样品的杉木凋落叶C/N比值达到试验要求。准确称取各样品所需的10±0.005 g经风干后的杉木凋落叶,装入网眼0.5 mm、大小为20 cm×20 cm的尼龙分解袋中,于2018年12月,分别放置于林下植被保留和林下植被去除共2种杉木人工林的3个预设样地内。放置时去除地表凋落物层,每个分解袋之间间隔50 cm,贴上标签,用竹钉固定四角在地表,保证分解袋紧贴表层土壤,让凋落叶自然分解。每种处理分解袋纵向放置,处理间设置3 m的缓冲带。
分解袋放置60 d、120 d、180 d、240 d和300 d后分别取样,每个样地每个处理取4袋,取样时戴一次性手套,每个分解袋均单独装入自封袋中,用于计算杉木凋落叶残留率。每次取样时,去除各分解袋上新鲜凋落物和林下植被去除林分中新生长的林下植被。取回的样品清除杂质,分别称质量后装入信封,做好标记,于烘箱中80 ℃烘干至恒重,然后称重,用粉碎机研磨后,用于营养元素含量测定。
2.3 测定方法
2.3.1 凋落叶养分含量测定
根据参考文献[18],凋落叶全氮和全碳采用全自动碳氮分析仪(Elemental Analyzer Vario ELIII,德国),全磷采用碱熔-钼锑抗比色法,全钾采用碱熔-火焰光度法,全钙和全镁含量采用EDTA络合滴定法,全锰含量测定采用原子吸收分光光度法。
2.3.2 凋落叶养分释放率的计算
养分释放率的计算公式为:

式中:Et为养分释放率(%),Wt为分解t时间后剩余的凋落叶干质量(g),Ct为凋落叶分解t时间后剩余的凋落叶养分含量(g·kg-1),Wo为凋落叶分解初始干质量(g),Co为凋落叶分解初始养分含量(g·kg-1)。
2.4 统计分析
初始数据采用Office 2019进行整理和统计,采用SPSS 26.0进行独立样本t检验、单因素方差分析、双因素方差分析、线性回归分析和相关分析,单因素分析选用Duncan差异显著性检验,相关性分析选用Person相关分析。
3 结果与分析
3.1 杉木凋落叶分解过程的C、N、P养分释放规律
各处理杉木凋落叶C养分含量在分解期间内均表现为养分释放,且不同时间和不同处理的C释放率存在显著差异(P<0.05)(见图1A)。就林分差异而言,2种林分杉木凋落叶C释放率之间无显著差异(P>0.05),但林下植被保留林分中杉木凋落叶C释放率整体上高于林下植被去除林分,说明林下植被保留有利于C的释放。随着分解时间的增加,碳含量因分解逐渐减少,从而造成C释放速率变缓。
如图1B所示,在试验期间内,杉木凋落叶N释放率均小于0,表明试验期间内2种林分杉木凋落叶N为养分富集。养分富集现象在凋落叶分解初期明显,林下植被保留林分中富集最明显主要是分解180 d时,而林下植被去除林分中在分解120 d时富集现象最明显,说明林下植被保留可以延缓N的富集现象。由多因素方差分析可知,相同时间的不同林分和不同处理杉木凋落叶N释放率无显著差异(P>0.05),而不同时间的N释放率达极显著差异(P<0.01)。
随着分解时间的增加,2种林分中P整体表现为养分释放,但林下植被保留林分中N3处理和林下植被去除林分中CK处理的杉木凋落叶P释放率均在分解240 d时小于0%,表现为养分富集(见图1C)。林下植被保留林分中,P释放率以N2处理较高,而林下植被去除林分中,不同分解时间段内各处理的P释放率变化较大,后期以N2处理较高。不同处理和不同时间的杉木凋落叶P释放率存在极显著差异(P<0.01),就林分差异而言,2种林分P释放率之间差异不显著(P>0.05)。
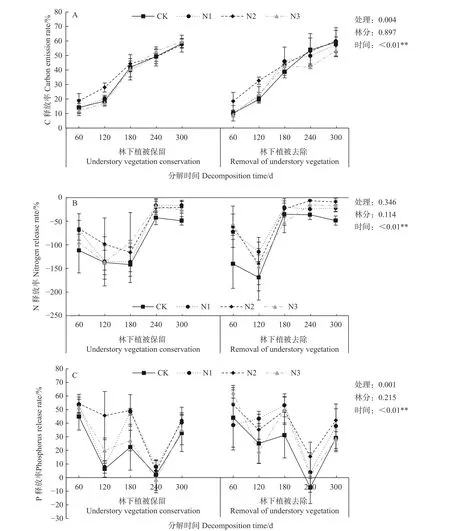
图1 杉木凋落叶分解过程C、N、P释放率(%)Fig.1 Release rate of C, N and P during C.lanceolata litter decomposition (%)
3.2 杉木凋落叶分解过程的K、Ca、Mg、Mn养分释放规律
在试验时间内,林下植被保留和林下植被去除2种林分中各处理的杉木凋落叶K、Ca、Mg均表现为养分释放,且3种元素释放率变化规律就林分而言均无显著差异(P>0.05)(见图2)。如图2A所示,K释放率随着分解时间的增加呈先降后升的变化趋势。林下植被保留林分中,各处理的K释放率均在分解180 d时达到最小值,而林下去除林分中,只有CK和N1处理在分解180 d时达到最小值,N2和N3处理的K释放率在分解120 d达到最小值。不同处理和不同时间的杉木凋落叶K释放速率存在极显著差异(P<0.01)。Ca和Mg释放率均随着分解时间的增加呈波浪变化趋势(见图2B、2C),但不同的是不同处理的Ca释放率存在显著差异(P<0.05),而不同处理和不同林分的Mg释放率之间不存在显著差异(P>0.05)。就Ca释放率而言,2种林分中各处理均在分解60 d时达到最大值,且凋落叶分解前期,林下植被去除林分较林下植被保留林分更有利于Ca的释放。如图2D所示,Mn大部分表现为养分释放,在分解181~300 d出现了养分富集现象,且不同处理的Mn释放率存在极显著差异(P<0.01),N1-N3处理的Mn释放率显著高于CK处理(P<0.05)。林下植被去除林分较林下植被保留林分更有利于杉木凋落叶中Mn的释放。
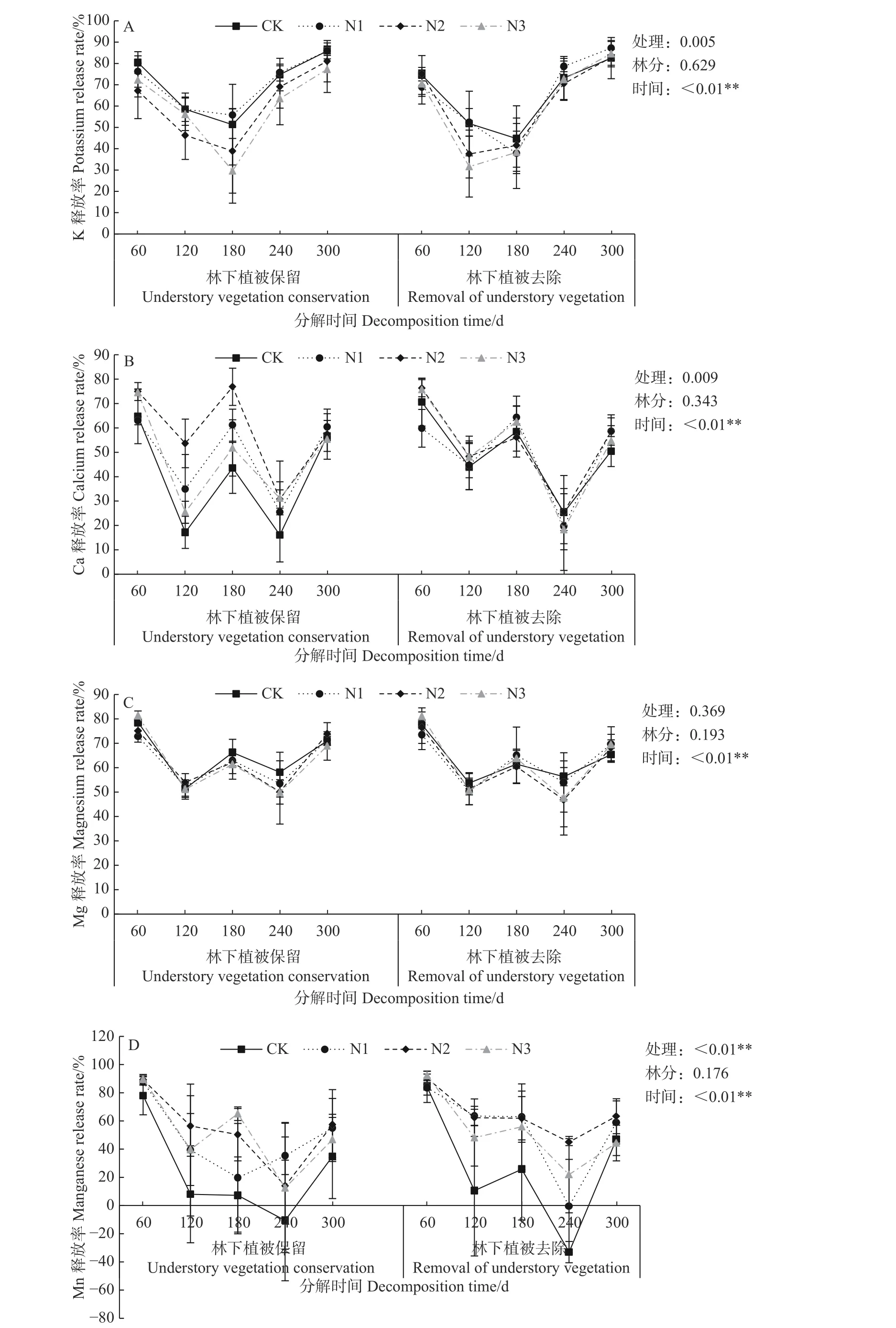
图2 杉木凋落叶分解过程K、Ca、Mg、Mn释放率(%)Fig.2 Release rate of K, Ca, Mg and Mn during C.lanceolata litter decomposition (%)
3.3 元素及元素比对分解过程的影响
由表2可知,N与林分、C/P呈显著(P<0.05)负相关,与C/N呈极显著(P<0.01)负相关;与K、Mn呈极显著(P<0.01)正相关,与N/P呈显著(P<0.05)正相关。C与时间显著(P<0.05)负相关,与阶段性失重、P、K、Mg、Mn均极显著(P<0.01)负相关;与C/N、C/P、N/P极显著(P<0.01)正相关。P与时间、阶段性失重、Ca、Mg、Mn呈极显著(P<0.01)正相关,与C/N、C/P、N/P均极显著(P<0.01)负相关。K与阶段性失重、Mg显著(P<0.05)正相关;与C/N呈极显著(P<0.01)负相关,与C/P呈显著(P<0.05)负相关。Ca与时间、Mg、Mn均极显著(P<0.01)正相关,与阶段性失重呈显著(P<0.05)正相关;与C/P、N/P均极显著(P<0.01)负相关。Mg与时间、Mn呈极显著(P<0.01)正相关,与阶段性失重呈显著(P<0.05)正相关;与C/N、C/P、N/P均极显著(P<0.01)负相关。Mn与时间、阶段性失重均极显著(P<0.01)正相关,与C/N、C/P、N/P均呈极显著(P<0.01)负相关。可见杉木凋落叶分解过程中自身各养分含量之间具有紧密的相关关系。

表2 凋落叶分解过程中各元素之间的Pearson相关系数Tab.2 Pearson correlation coefficient among various elements during litter decomposition
可见经外源氮施入处理后,N含量不再成为养分限制因子,杉木凋落叶分解过程主要受P含量、C/P比值和N/P比值所调控,P含量的提高有利于杉木凋落叶的分解,其中Mn含量在一定范围内对于杉木凋落叶的分解具有重要作用。
4 结论与讨论
森林凋落物的分解过程包括淋溶、自然粉碎和代谢作用等物理和化学过程[19]。在大区域范围内,气候是凋落物分解的主要调控因素;而底物质量则主要影响小区域范围内凋落物的分解[20],其中影响凋落物分解的主要化学因素是基质碳形态,碳氮相互作用对于凋落物分解过程也有尤为重要的影响。杉木凋落物质量主要是影响凋落物分解环境中的微生物群落,其养分含量越高(如C/N和C/P比),微生物群落生长加快,从而间接影响到凋落物的分解速率[19],而且有研究表明导致分解周期增长、分解周转较慢的重要原因是N含量的贫乏[13]。人类活动造成全球氮沉降的不断加剧,N作为影响杉木凋落物分解的重要因素,在杉木人工林生态系统中的有效性有所提升,对凋落物分解的内在机理仍需进一步研究。本研究,杉木凋落叶在分解300 d过程中,总体表现为试验前期和试验后期分解较慢,试验中期分解较快,与林开敏等[21]和何宗明等[22]对杉木凋落物分解研究结果有所差异,这可能是由于本试验时间较短,并未观测到杉木凋落叶的完整分解周期。本试验结果表明,外源氮的输入对于杉木凋落叶的分解具有两重性,即低促高抑的作用,且随着分解时间的增长,促进作用逐渐减弱,抑制作用逐渐增强。在试验期间内,只有杉木凋落叶C浓度低于初始浓度,其余元素浓度普遍高于杉木凋落叶初始浓度。不同处理的杉木凋落叶的Mn、Ca、P浓度差异显著,且以CK处理的浓度最高,说明高C/N有利于Mn、Ca、P含量的积累。
只有N在整个杉木凋落叶分解过程中呈现富集现象,说明N对于分解者有养分供应的关系,相关性分析表明杉木凋落叶中N与阶段性失重呈负相关,且外源氮的输入缓解了分解者对N的需求。已有研究证明,分解初期N含量是分解控制的主要因素之一,而后期以木质素为主导[23]。外源氮的输入使得N不再成为主要的限制因素,研究结果表明C浓度和C/N比值出现先下降后上升的变化趋势。自1930年Melin[24]首次提出用C/N值来分析凋落叶的特征以来,已有大量研究对凋落物分解的C/N临界值进行了探索。大量研究表明C/N值范围被确定在20∶1~30∶1[25-27],而本研究结果,C/N临界值范围在17∶1~22∶1,这可能是由于林分对微生物的影响,从而导致该值偏小,结果有待进一步验证。
杉木凋落叶P浓度随着分解时间的增长总体上呈上升趋势,但C/P值不断下降,且相关性分析表明P与阶段性失重呈极显著正相关,而与P相关的指标,如C/P比和N/P等指标,与阶段性失重具有极显著负相关关系。可见相比于N而言,本研究中P更可能是调控杉木凋落叶分解的重要指标之一。已有研究证明当N/P比值>16时,P会限制凋落物的分解[28],且C/P比值范围大于100∶1~480∶1时也会限制凋落物的分解[25,29,30]。本研究中,杉木凋落叶N/P比值范围为29∶1~119∶1,C/P比值范围为643∶1~3 545∶1,远高于以上研究的临界值。这可能是由于本研究地区位于亚热带,土壤存在风蚀现象,导致P的相对消耗,且分解初期杉木凋落叶P含量较低,不能满足分解者生长的需求,从而分解者需要通过摄取外界环境中的P来满足生长的需求,故造成杉木凋落叶在分解过程中出现P富集现象。相关性分析表明P与Ca、Mg、Mn等元素均具有极显著的相关性,可以推测出,N充足时,P成为调控杉木凋落叶分解的重要指标。
已有大量研究发现,N和P在凋落物分解过程中出现了富集现象[19,31,32],本研究Mn也在分解过程中出现了富集现象,且K、Ca、Mg、Mn等元素与阶段性失重有显著和极显著正相关性。就养分释放规律而言,在分解前60 d养分释放速率普遍较高,这可能是由于受淋溶作用的影响,导致凋落叶中可溶性碳水化合物快速损失。总体上,在整个分解期间,以K、Ca等金属元素释放率较高,这与K主要以可溶性盐基的形式存在于凋落物内,但Ca释放速率波动较大,而Mg释放速率相对较低,易被淋溶,与大量研究结果基本一致[33]。元素的转移方式决定了其丢失速度[34],从元素动态模型来看,杉木凋落叶元素释放规律与失重率的动态变化具有趋同效应,杉木凋落物具有元素本身释放与自身质量降低的双重释放格局,这与Ribeiro等[35]的研究结果相似。值得注意的是,微量元素与凋落物分解的相关性,进而对限制凋落物分解的化学元素进行重要性排序。由相关性分析可知,杉木凋落叶中仅N和C/N与林分存在显著相关性,林分差异对于其余元素无显著影响。
猜你喜欢
天津医科大学学报(2021年1期)2021-12-05
防护林科技(2020年6期)2020-08-12
农民致富之友(2020年8期)2020-05-11
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
读写算(2018年6期)2018-08-22
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
绿色科技(2016年15期)2016-10-11
考试周刊(2016年63期)2016-08-15
科技视界(2016年1期)2016-03-30
