
毛蕊异黄酮对人骨关节炎软骨细胞增殖和凋亡的影响*
2021-12-20 05:36廖军锋吕晓宇
广西医科大学学报 2021年11期
廖军锋,王 虹,吕晓宇
(中国人民解放军南部战区总医院康复医学科,广州 510010)
骨关节炎是一种多发生于中、老年人的炎症性关节病,其发病机制尚未明确,且缺乏有效的治疗手段[1]。研究显示,骨关节炎软骨细胞凋亡是骨关节炎发生的关键[2],抑制软骨细胞凋亡是骨关节炎非手术治疗药物开发的研究热点。毛蕊异黄酮是从中药黄芪中提取的单体成分,具有多种药理活性。研究显示,毛蕊异黄酮可通过激活PI3K/Akt和Nrf2/HO-1信号通路减轻缺糖缺氧诱导的心肌细胞损伤[3];可通过降低脑组织炎症反应和氧化应激减轻脑缺血再灌注损伤大鼠脑组织损伤[4],表明毛蕊异黄酮具有良好的抗炎抗氧化作用,但目前关于毛蕊异黄酮对骨关节炎软骨细胞损伤的影响和机制还未见相关报道。miR-502-5p属于微小RNA(miRNA)家族成员,参与增殖、分化和凋亡等生理或病理过程,与肿瘤等多种疾病的发生发展密切相关[5-6]。研究显示,骨关节炎患者软骨组织中miR-502-5p的表达明显降低,上调miR-502-5p 可靶向负调控TRAF2的表达抑制IL-1β诱导的软骨细胞凋亡和炎症反应,miR-502-5p可作为减轻软骨细胞损伤的分子靶点[7]。因此,本研究通过体外分离培养骨关节炎软骨细胞,探究毛蕊异黄酮对骨关节炎软骨细胞增殖和凋亡的影响及其能否调控miR-502-5p 表达发挥作用,以期为其用于骨关节炎的治疗提供一定的实验依据。
1 资料与方法
1.1 临床资料
选择2018 年2 月至2019 年3 月中国人民解放军南部战区总医院康复医学科收治的10 例行全膝关节置换术的骨关节炎患者为研究对象,其中男4例,女6例,平均(60.28±6.14)岁。取材位置为股骨髁内、外髁负重过度区的关节软骨组织,液氮保存。样本采集经本院伦理委员会批准,患者均签署知情同意书。
1.2 实验试剂
毛蕊异黄酮,纯度≥98%,上海贤鼎生物科技有限公司;阿司匹林,片剂,规格0.3 g,远大医药(中国)有限公司;DMEM 培养基、细胞周期检测试剂盒、Annexin V-FITC/PI 细胞凋亡试剂盒和二喹啉甲酸(BCA)蛋白检测试剂盒,北京索莱宝;LipofectamineTM2000 试剂盒,美国Invitrogen 公司;胎牛血清(FBS),浙江天杭生物科技公司;anti-miR-502-5p、anti-miR-NC和PCR引物,上海生工;B淋巴细胞瘤-2(Bcl-2)、B淋巴细胞瘤-2相关蛋白(Bax)和甘油醛-3-磷酸脱氢酶(GAPDH)单克隆抗体,美国Santa Cruz 公司;miRNA 提取试剂盒、逆转录试剂盒和PCR试剂盒,大连宝生物。
1.3 方法
1.3.1 骨关节炎软骨细胞分离培养和转染 在无菌条件下,将骨关节炎软骨组织剪成1 mm×1 mm×1 mm,加入0.2 %Ⅱ胶原酶消化30 min,加含10 %FBS 的DMEM 培养基终止消化,过200 目筛,滤液1 000 r/min 离心5 min。弃上清,用含10 % FBS 的DMEM 培养基重悬细胞,接种至培养瓶中,于培养箱中培养。5 d后,更换新鲜培养基,此后每2~3 d更换1 次培养基。待细胞密度至80%时,加0.25%胰蛋白酶消化,传代培养。将对数期骨关节炎软骨细胞接种于6 孔板中(5.0×105个/孔),用LipofectamineTM2000 脂质体法,分别转染anti-miR-NC、antimiR-502-5p。转染6 h 后,更换培养基。再培养24 h,收集细胞备用。
1.3.2 细胞分组处理 将未转染的骨关节炎细胞分为对照组、毛蕊异黄酮低剂量组、毛蕊异黄酮中剂量组、毛蕊异黄酮高剂量组和阳性对照组,其中对照组细胞用常规培养基培养,毛蕊异黄酮低、中、高剂量组细胞分别用含0.1 mg/L、1 mg/L、10 mg/L毛蕊异黄酮的培养基培养,阳性对照组细胞用含9 mg/mL 阿司匹林的培养基培养。转染anti-miRNC、anti-miR-502-5p 的细胞用含10 mg/L 毛蕊异黄酮的培养基培养,分别记为毛蕊异黄酮高剂量+antimiR-NC 组、毛蕊异黄酮高剂量+anti-miR-502-5p组。
1.3.3 细胞计数试剂盒8(CCK-8)法检测细胞增殖试验 将未转染及转染anti-miR-NC、anti-miR-502-5p 的骨关节炎细胞均接种于96 孔板中(1.0×104个/孔),培养12 h 后,弃培养基,按照上述“1.3.2 项”分组处理。分别培养24 h、48 h 和72 h 后,加10 μL CCK-8 试剂。孵育2 h 后,酶标仪450 nm 测光密度(OD)值。
1.3.4 流式细胞仪检测细胞周期 将未转染及转染anti-miR-NC、anti-miR-502-5p的骨关节炎细胞均接种于24孔板中(2.5×104个/孔),培养12 h后,弃培养基,按照上述“1.3.2 项”分组处理。培养48 h 后,收集细胞,并使用PBS 清洗2 次。加1 mL 预冷的70%乙醇,混悬细胞,4 ℃冰箱中固定12 h,1 000 r/min离心5 min,弃上清,并用PBS清洗细胞2次。加0.5 μL PI,37 ℃孵育30 min,上流式细胞仪检测。
1.3.5 流式细胞仪检测细胞凋亡 细胞培养和分组同“1.3.4项”,培养结束后,收集细胞,用PBS清洗2 次。加500 μL 结合缓冲液,重悬细胞。依次加10 μL Annexin V-FITC 和5 μL PI,避光孵育15 min后,上流式细胞仪检测细胞凋亡。
1.3.6 Western blotting 法检测Bax 和Bcl-2 蛋白表达 细胞培养和分组同“1.3.4 项”,培养结束后,收集细胞,用RIPA 试剂提取细胞中总蛋白。经BCA法定量、电泳、转膜和封闭1 h后,分别于Bax(1∶500)、Bcl-2(1∶500)和GAPDH(1∶800)一抗孵育液中4 ℃冰箱过夜。再于山羊抗兔二抗(1∶3 000)孵育液中37 ℃孵育1 h。最后加显影液避光显影,曝光拍照。
1.3.7 实时荧光定量PCR(RT-qPCR)检测细胞中miR-502-5p表达 细胞培养和分组同“1.3.4项”,培养结束后,收集细胞,用miRNA 提取试剂盒细胞中总RNA,逆转录为cDNA 后,行PCR 扩增。反应条件:95 ℃5 min,95 ℃10 s,60 ℃30 s,72 ℃30 s,共35 个循环。引物序列如下,miR-502-5p 上游:5'-CGGGCATCCTTGCTATCTG-3',下游:5'-CAGCCACAAAAGAGCACAAT-3';内参U6 上游:5'-CTCGCTTCGGCAGCACA-3',下游:5'-AACGCTTCACGAATTTGCGT-3'。采用2-△△Ct法计算miR-502-5p相对于内参U6的表达水平。
1.4 统计学方法
采用SPSS 22.0 软件对数据进行统计学分析。计量资料以均数±标准差()表示,两组间比较用独立样本t检验,多组间比较用单因素方差分析,组间两两比较用SNK-q检验。以P<0.05为差异有统计学意义。
2 结果
2.1 毛蕊异黄酮对骨关节炎软骨细胞增殖的影响
对分离培养的不同传代的OA软骨细胞形态学进行观察,发现第一代呈上皮样,会有高亮圆形细胞,随着传代逐渐减少,至第4~6代细胞呈现梭形生长,见图1。与对照组比较,毛蕊异黄酮中、高剂量组骨关节炎软骨细胞增殖活性升高(均P<0.05),细胞周期G0~G1 期缩短(P<0.05),而S 期延长,并呈剂量依赖性(P<0.05),G2~M 期无明显变化(均P>0.05)。与对照组比较,阳性对照组骨关节炎软骨细胞增殖活性升高(P<0.05),细胞周期G0~G1期缩短(P<0.05),而S 期延长(P<0.05),G2~M 期无明显变化(P>0.05)。与阳性对照组比较,毛蕊异黄酮高剂量组骨关节炎软骨细胞增殖活性及细胞周期均无明显变化(均P>0.05),见表1。

图1 不同传代的OA软骨细胞形态学变化(×200)
表1 毛蕊异黄酮对骨关节炎软骨细胞细胞细胞增殖活性和细胞周期的影响,n=3
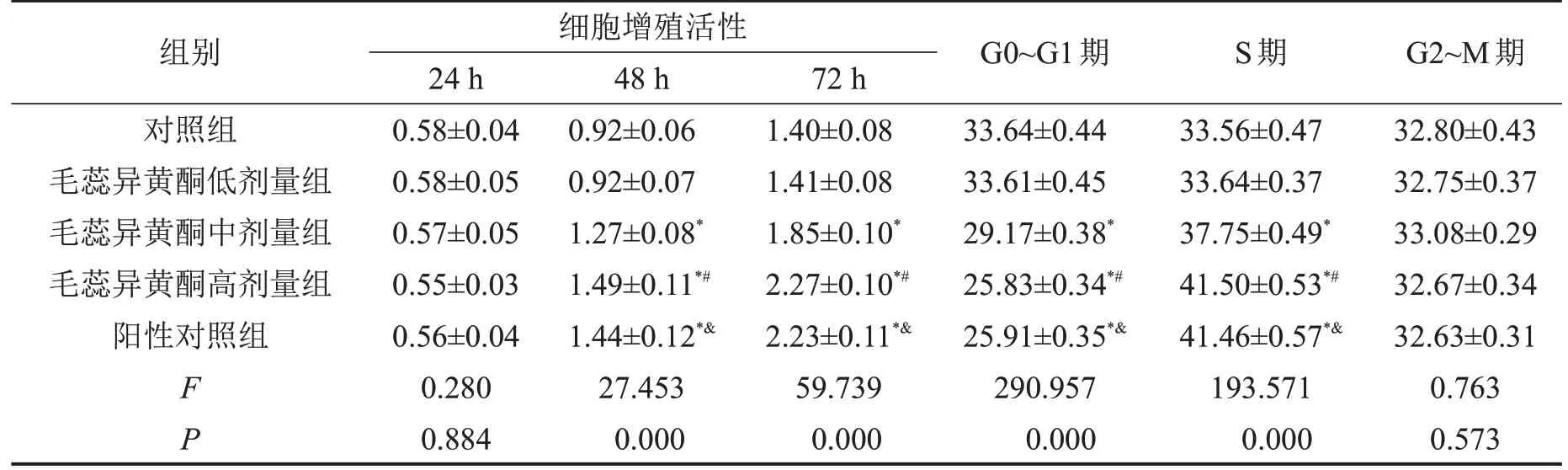
表1 毛蕊异黄酮对骨关节炎软骨细胞细胞细胞增殖活性和细胞周期的影响,n=3
与对照组相比,*P<0.05;与毛蕊异黄酮低剂量组相比,#P<0.05;与毛蕊异黄酮中剂量组相比,&P<0.05。
2.2 毛蕊异黄酮对骨关节炎软骨细胞凋亡的影响
与对照组比较,毛蕊异黄酮中、高剂量组骨关节炎软骨细胞凋亡率和Bax蛋白表达降低,而Bcl-2蛋白表达升高(均P<0.05),并呈一定剂量依赖性。与对照组比较,阳性对照组骨关节炎软骨细胞凋亡率和Bax 蛋白表达降低(均P<0.05),而Bcl-2蛋白表达升高(均P<0.05)。与阳性对照组比较,毛蕊异黄酮高剂量组骨关节炎软骨细胞凋亡率及细胞中Bax和Bcl-2蛋白表达均无明显变化(均P>0.05),见图2、图3和表2。
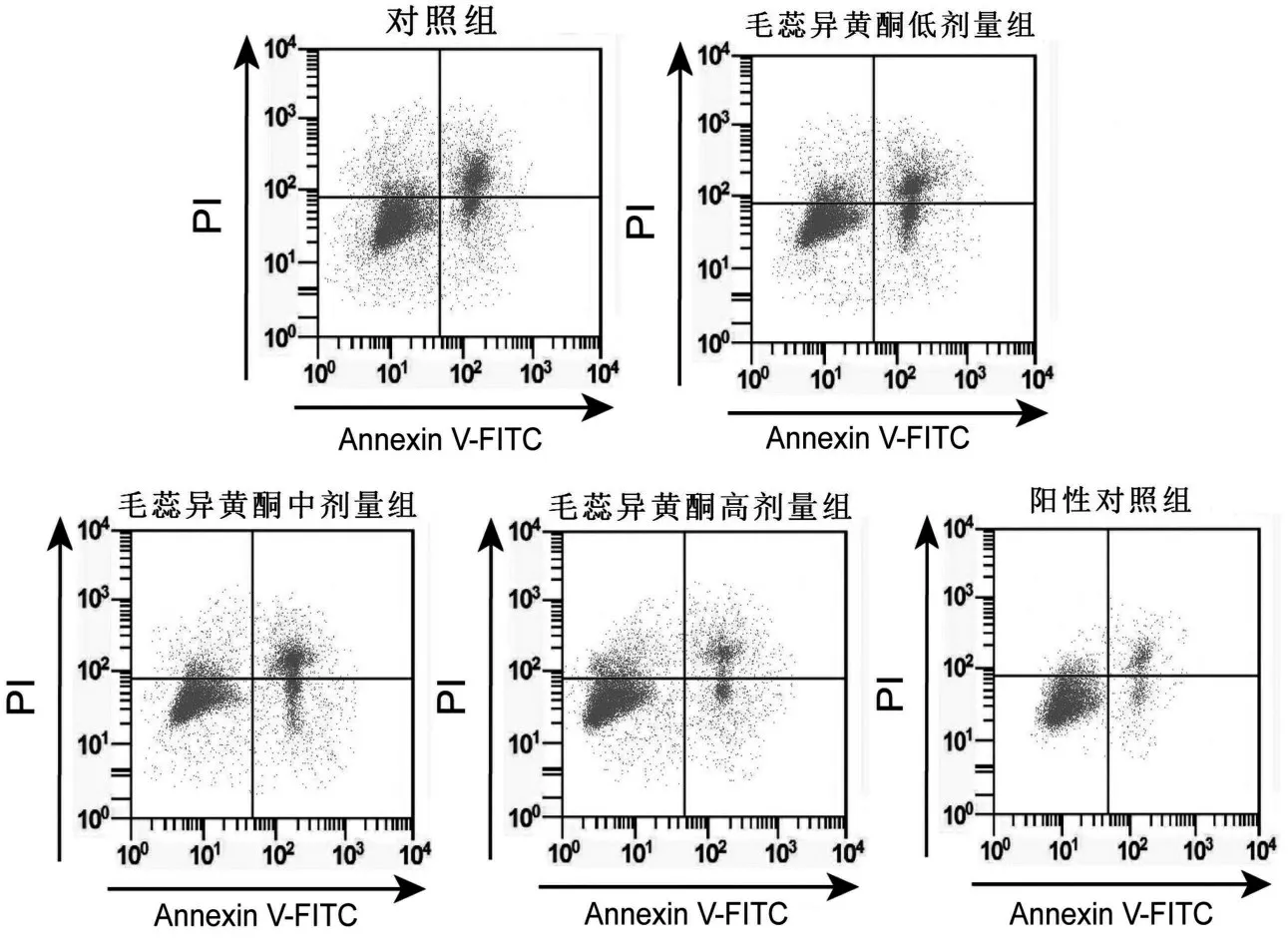
图2 毛蕊异黄酮对骨关节炎软骨细胞凋亡的影响
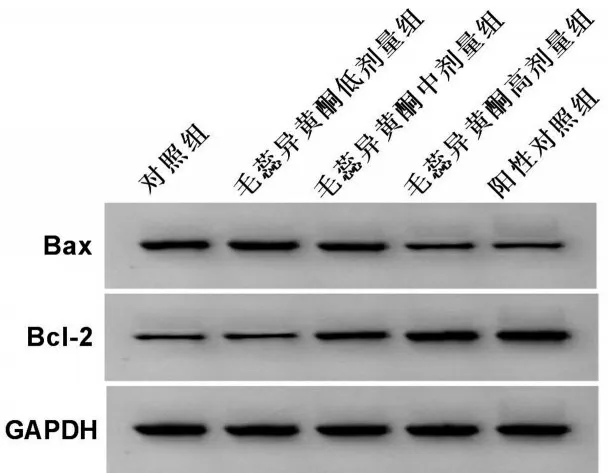
图3 毛蕊异黄酮对骨关节炎软骨细胞中Bax、Bcl-2蛋白表达的影响
表2 毛蕊异黄酮对骨关节炎软骨细胞凋亡及凋亡蛋白Bax、Bcl-2表达的影响,n=3
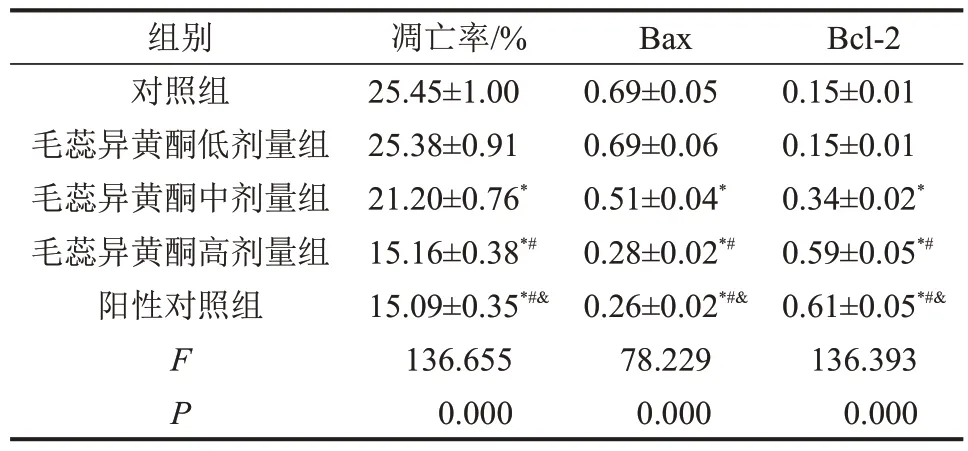
表2 毛蕊异黄酮对骨关节炎软骨细胞凋亡及凋亡蛋白Bax、Bcl-2表达的影响,n=3
与对照组相比,*P<0.05;与毛蕊异黄酮低剂量组相比,#P<0.05;与毛蕊异黄酮中剂量组相比,&P<0.05。
2.3 毛蕊异黄酮对骨关节炎软骨细胞中miR-502-5p表达的影响
与对照组比较,毛蕊异黄酮中、高剂量组骨关节炎软骨细胞中miR-502-5p 表达升高(P<0.05),并呈剂量依赖的趋势。与对照组比较,阳性对照组骨关节炎软骨细胞中miR-502-5p 表达升高(P<0.05)。与阳性对照组比较,毛蕊异黄酮高剂量组组骨关节炎软骨细胞中miR-502-5p 表达无显著变化(P>0.05),见表3。
表3 毛蕊异黄酮对骨关节炎软骨细胞中miR-502-5p 表达的影响,n=3
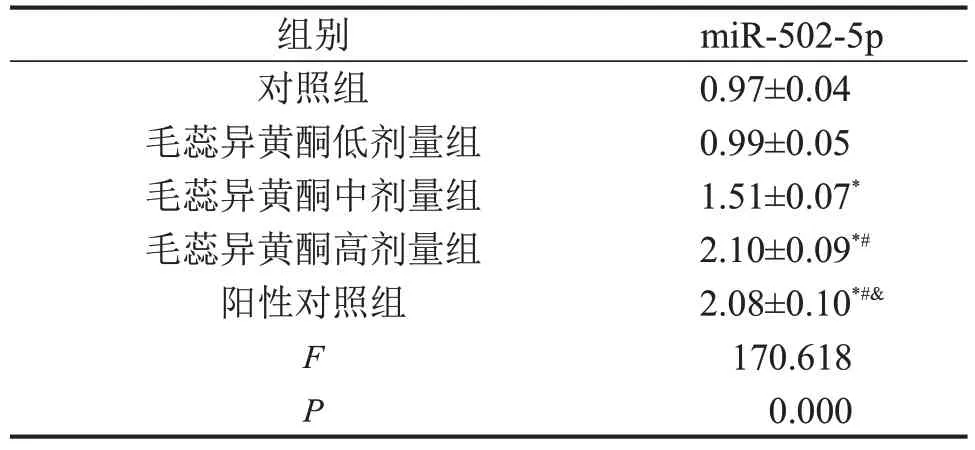
表3 毛蕊异黄酮对骨关节炎软骨细胞中miR-502-5p 表达的影响,n=3
与对照组相比,*P<0.05;与毛蕊异黄酮低剂量组相比,#P<0.05;与毛蕊异黄酮中剂量组相比,&P<0.05。
2.4 下调miR-502-5p 对毛蕊异黄酮处理的骨关节炎软骨细胞增殖的影响
与毛蕊异黄酮高剂量+anti-miR-NC 组比较,毛蕊异黄酮高剂量+anti-miR-502-5p 组骨关节炎软骨细胞增殖活性降低(P<0.05),细胞周期G0~G1 期延长(P<0.05),而S 期缩短(P<0.05),G2~M 期无显著变化(P>0.05),见表4。
表4 下调miR-502-5p对毛蕊异黄酮处理的骨关节炎软骨细胞增殖的影响,n=3
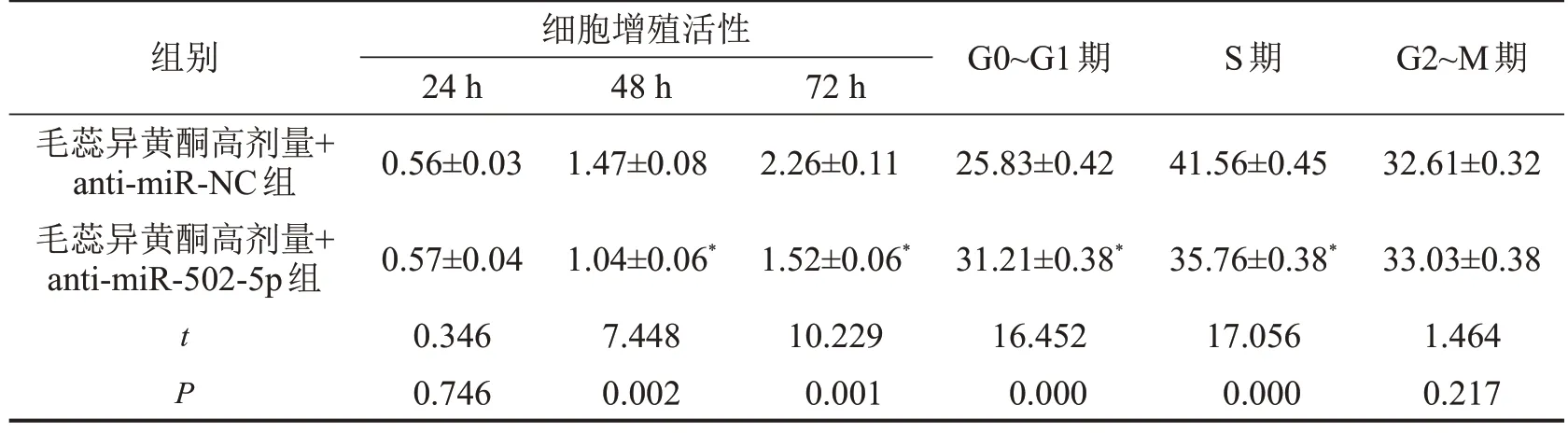
表4 下调miR-502-5p对毛蕊异黄酮处理的骨关节炎软骨细胞增殖的影响,n=3
与毛蕊异黄酮高剂量+anti-miR-NC组相比,*P<0.05。
2.5 下调miR-502-5p 对毛蕊异黄酮处理的骨关节炎软骨细胞凋亡的影响
与毛蕊异黄酮高剂量+anti-miR-NC 组比较,毛蕊异黄酮高剂量+anti-miR-502-5p 组骨关节炎软骨细胞凋亡率和Bax 蛋白表达升高(均P<0.05),而Bcl-2蛋白表达降低(P<0.05),见图4、图5和表5。
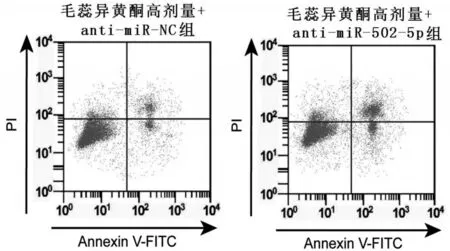
图4 下调miR-502-5p 对毛蕊异黄酮处理的骨关节炎软骨细胞凋亡的影响
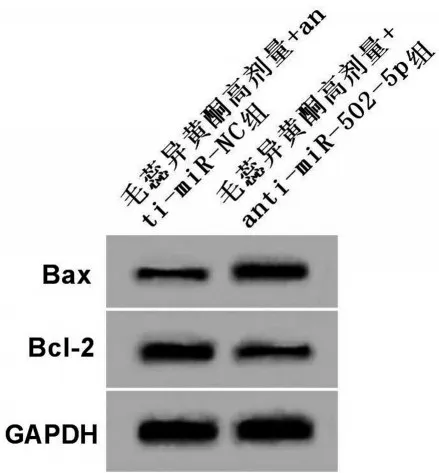
图5 下调miR-502-5p 对毛蕊异黄酮处理的骨关节炎软骨细胞Bax和Bcl-2蛋白表达的影响
表5 下调miR-502-5p对毛蕊异黄酮处理的骨关节炎软骨细胞凋亡及Bax和Bcl-2蛋白表达的影响,n=3
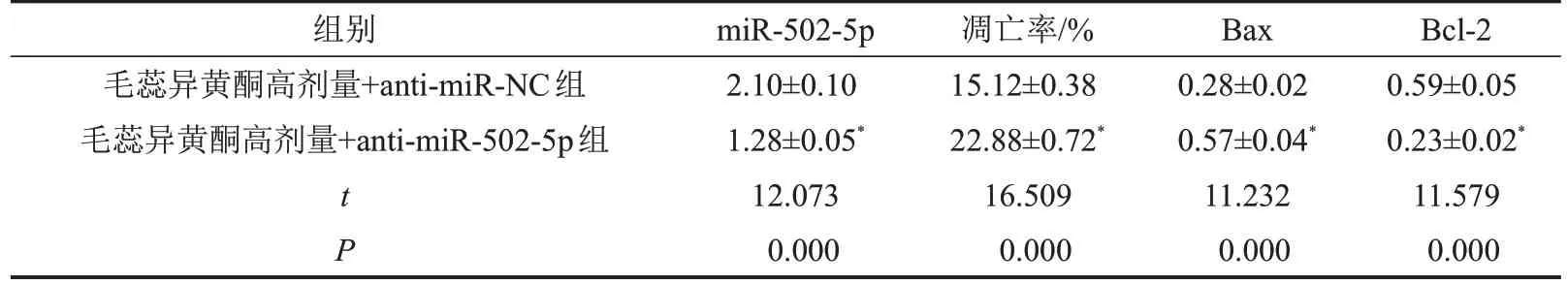
表5 下调miR-502-5p对毛蕊异黄酮处理的骨关节炎软骨细胞凋亡及Bax和Bcl-2蛋白表达的影响,n=3
与毛蕊异黄酮高剂量+anti-miR-NC组相比,*P<0.05。
3 讨论
软骨细胞是关节软骨中的唯一细胞,参与维持软骨组织结构和功能稳定。骨关节炎患者软骨细胞过度凋亡,进而引发软骨基质减少,抑制骨关节炎软骨细胞细胞凋亡是骨关节炎治疗的途径之一。毛蕊异黄酮为中药黄芪的主要活性成分之一,具有多种药理活性。研究显示,毛蕊异黄酮可提高缺氧缺糖/复氧复糖诱导的心肌细胞活力,并抑制细胞凋亡,减轻细胞损伤[8];毛蕊异黄酮可能通过提高内源抗氧化酶T-SOD 和GSH-Px 活性、降低脂质过氧化损伤及脑组织AchE活性来改善D-半乳糖致衰老小鼠的学习记忆能力[9];毛蕊异黄酮可能通过激活Nrf2 信号通路缓解了大鼠蛛网膜下腔出血后脑水肿和血脑屏障破坏,减轻了神经炎症反应和细胞凋亡[10]。
本研究显示,毛蕊异黄酮可提高骨关节炎软骨细胞增殖活性,阻滞了其细胞周期进程,降低了细胞凋亡率,并具有一定的剂量依赖性(P<0.05),说明毛蕊异黄酮可促进骨关节炎软骨细胞增殖,且抑制软骨细胞凋亡。细胞凋亡是一种受多种分子调控的程序性死亡过程。Bax 和Bcl-2 参与调控细胞凋亡,其中Bax为促凋亡蛋白,其表达增加时形成同源二聚体,引起线粒体膜通透性改变,细胞色素C释放量增加,进而激活caspases级联反应,诱导细胞凋亡[11]。而Bcl-2是一种抗凋亡分子,其表达增加时则与Bax 形成异源二聚体,减弱Bax 的促凋亡作用。本研究显示,一定剂量的毛蕊异黄酮可抑制骨关节炎软骨细胞中Bax的蛋白表达,而促进了Bcl-2蛋白表达(P<0.05),进一步说明毛蕊异黄酮抑制了骨关节炎软骨细胞凋亡,具有治疗骨关节炎的作用。
骨关节炎发病率呈增长趋势,探究其发病机制对治疗药物的研发具有重要现实意义。研究显示,真核生物中存在大量miRNA,这些miRNA 参与调控细胞增殖和凋亡等生命活动,可作为包括骨关节炎在内的多种疾病的治疗靶点[12-14]。研究已表明,多种miRNA在骨关节炎中异常表达,在骨关节炎发生发展中起重要作用。例如,miR-145 在骨关节炎软骨细胞中呈低表达,上调其表达可靶向抑制BNIP3和调节Notch信号通路来减少骨关节炎软骨细胞凋亡[15];骨关节炎软骨组织和细胞中miR-124呈低表达,上调其表达可抑制软骨细胞凋亡[16]。miR-502-5p 在滑膜组织中的表达与滑膜增生及炎症反应密切相关,通络止痛凝胶制剂可能通过调控miR-502-5p 等基因的表达减轻膝骨关节炎滑膜增生及炎症反应[17]。本研究显示,毛蕊异黄酮可上调骨关节炎软骨细胞中miR-502-5p 的表达,而下调miR-502-5p 则逆转了毛蕊异黄酮对骨关节炎软骨细胞增殖的促进作用及抑制凋亡作用,提示毛蕊异黄酮可能通过上调miR-502-5p 表达来影响骨关节炎软骨细胞增殖和凋亡。
综上,毛蕊异黄酮可增强骨关节炎软骨细胞的增殖能力,并减少细胞凋亡,具有治疗骨关节炎的潜在价值,其作用机制可能与上调细胞中miR-502-5p表达有关。
猜你喜欢
中华骨与关节外科杂志(2022年1期)2022-08-31
现代临床医学(2022年2期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年2期)2022-03-29
食品与健康(2020年4期)2020-05-18
家庭医学(2019年6期)2019-06-27
中国食品(2018年7期)2018-09-10
家庭医药·快乐养生(2017年4期)2017-04-19
家庭百事通·健康一点通(2017年2期)2017-02-23
家庭医药(2016年1期)2016-01-20
