
不同氮沉降对耐冬山茶和茶梅生长及生理特性的影响
2021-12-14 08:15:50马翔李辉郭霄李明燕柳健李有刘庆超王奎玲刘庆华
青岛农业大学学报(自然科学版) 2021年4期
马翔,李辉,郭霄,李明燕,柳健, 李有,刘庆超,王奎玲,刘庆华
(1.青岛农业大学园林与林学院,山东青岛 266109;2. 济南市林场,山东济南 250100)
人类社会的发展与进步将会使得全球范围内的活性氮排量持续增加[1-3],氮的排放和沉积率到2050年将增加一倍,全球氮沉降量将达到1.9×108t·a-1。氮沉降增加一是使土壤内的氮含量增加[4];二是由于降水的不均衡性,导致了不同地区的氮素资源异质性[5]。氮沉降对植物的影响具有两面性,一方面氮沉降有利于植物的生长发育[4],保证了植物生长发育所需的氮素营养物质的充足,促进植物光合作用,提高生产力[1, 6-8];另一方面,氮元素过多会抑制植物的生长发育和各项生理活动,例如破坏植物细胞的膜组织,造成植物养分失衡,高渗透压会导致植物失水过多,甚至会使植株干枯死亡[3-4, 9]。资源异质性会使生态系统中各部分获取的资源变得不均衡,原有的细胞结构会受到破坏,降低生物量和生物多样性。目前已有大量围绕养分空间异质性而展开的试验研究,有的研究表明养分空间异质性不仅能影响生物的总生物量,还会影响生物对生物量的分配[10]。
耐冬山茶[Camelliajaponica(NaiDong)],山茶科山茶属常绿阔叶灌木或小乔木,是分布于胶东半岛地区的一个山茶原生种群[4],因其花在最严寒时节绽放,被当地称之为“耐冬山茶”。茶梅(Camelliasasanqua),山茶科山茶属常绿灌木或小乔木,主要分布于中国和日本西南部,花芳香,花期长,花期自10月下旬至来年4月,是山茶属中四大观赏栽培类群之一[11]。耐冬山茶和茶梅因具有极高的观赏价值而被广泛应用于园林绿化当中。前人对耐冬山茶和茶梅的研究大多集中在种质资源、品种分类、杂交育种等方面,关于氮沉降和时间异质性的研究较少。本文以耐冬山茶和茶梅为研究对象,分析不同氮沉降胁迫下耐冬山茶和茶梅的生理生态学适应机制,为今后耐冬山茶和茶梅的资源开发利用提供理论支持。
1 材料与方法
1.1 试验材料
研究对象为1年生耐冬山茶和茶梅实生苗。7月于青岛市植物园采集种子,12月至来年3月将种子沙藏催芽,3月初种植于花盆(高15 cm、直径15 cm)中,栽培基质是经过充分混合的相同体积的草炭和沙土。6月初,对长势正常的幼苗进行筛选,耐冬山茶幼苗和茶梅幼苗各自筛选出63盆,每7盆为1组。
1.2 试验设计
2018年夏季在青岛农业大学温室内进行试验。设置3个氮沉降量和3个氮沉降添加频率,3个氮沉降量分别为N0对照组(0 g·m-2·a-1)、N1轻度氮沉降(6 g·m-2·a-1)、N2重度氮沉降(12 g·m-2·a-1)[4];3个不同氮沉降量使用的添加液的成分和浓度见表1。3个氮沉降添加频率分别为F1高添加频率(6 d添加一次)、F2中添加频率(12 d添加一次)、F3低添加频率(18 d添加一次)。耐冬山茶幼苗和茶梅幼苗各9个处理组,每组7个重复,试验处理时间共计108 d。整个试验过程中,保证室温均衡一致且通风良好,及时进行正常的养护管理工作,防止夏季经常出现的多风多雨天气的影响,避免杂草、病虫害等发生。

表1 3个不同氮沉降量使用的添加液的成分和浓度表Table 1 Composition and concentration table of additive solutions used at three different nitrogen settlement
1.3 试验指标的测定[12]
1.3.1 生长指标的测定
在最后一次添加氮沉降处理溶液的7 d后对耐冬山茶幼苗和茶梅幼苗的株高(H)、基径(BD)和冠面积(SCA)等生长指标进行测量。冠面积采用Li等[2]所述的公式测量并计算。
SCA=a×b×0.25π
其中:a为每株幼苗最大伸展处测量所得冠幅长,cm;b为在其平面垂直方向测量所得冠幅宽,cm。
比叶面积测定:每株植株摘取完全展开的健康成熟叶片1片,用便携式叶面积仪Yaxin-1241(北京雅欣理仪科技有限公司)测量叶片面积(SLA)并记录。最后将叶片放入烘箱中用85 ℃烘48 h使其完全脱水后称量其干质量(MLDW)。根据公式计算比叶面积(SLA):
SLA=SLA/MLDW× 100%[2]
生物量的测定:将试验所用的全部的耐冬山茶幼苗和茶梅幼苗整株挖出,切割分解为根、茎、叶三部分,分别放入烘箱中80 ℃烘48 h,最后使用电子天平各自称量根、茎、叶三部分的质量并记录,根据以下公式计算生物量的分配[3]:
总生物量MTB=MRB+MSB+MLB;
根生物量比(RMR)=MRB/MTB;
叶生物量比(LMR)=MLB/MTB;
根冠比(RS)=MRB/ (MLB+MSB)
其中:MRB根生物量,MTB总生物量,MLB叶生物量,MSB茎生物量。
1.3.2 生理指标的测定
气体交换参数测定:使用便携式光合仪CIRAS-3(汉莎科学仪器有限公司,UK)对植株叶片的净光合速率(Pn)、细胞间隙CO2浓度(Ci)、蒸腾速率(E)、气孔导度(Gs)等值进行测量,测量时的参数设置为光强 1 000 μmol·m-2·s-1,叶室温度 27 ℃,湿度为 60%, CO2浓度为 400 μmol·mol-1。并根据公式计算气孔限制值(LS):
LS=1-Ci /Ca,(Ca为空气CO2浓度),
叶绿素荧光参数测定:使用植物效率分析仪Pocket PEA(汉莎科学仪器有限公司,UK)对植株叶片的叶绿素荧光参数进行测量[12]。
1.4 数据处理
用Excel 2019整理数据,釆用SPSS 21对数据进行正态检验和方差齐性检验后进行单因素和双因素的ANOVA分析,比较各个参数分别在不同氮沉降量和氮沉降添加频率条件下的差异,以及两因素间是否存在交互作用。存在显著差异之后进行邓肯(Duncan)多重比较(α=0.05),并用字母进行标记。
2 结果分析
2.1 耐冬山茶和茶梅的生长指标对不同氮沉降的响应
由表2可以看出,氮沉降和添加频率及二者的交互作用,对耐冬山茶的生长指标无影响,而对茶梅的冠面积影响极显著。由图1可知,茶梅的株高在重度氮沉降和低添加频率的条件下达到最大值(9.57 cm),相较于相同氮沉降水平下的高添加频率和中添加频率,分别提高了15.7%和7.5%;氮沉降对茶梅基径的影响不显著;茶梅冠面积受氮沉降的影响显著,加氮组的茶梅冠面积低于对照组。
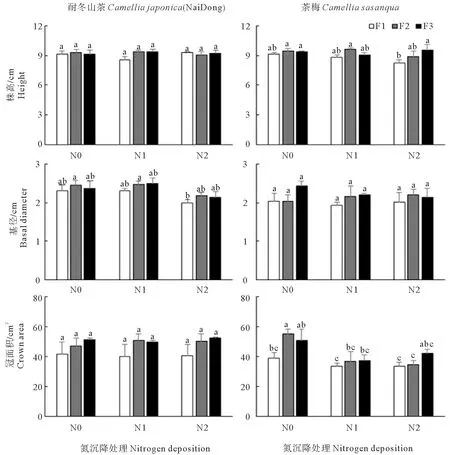
图1 氮沉降量和氮沉降添加频率的改变对植株生长指标的影响Fig.1 Effects of nitrogen deposition rate and nitrogen deposition frequency on growth parameter of plant

表2 氮沉降量和添加频率对耐冬山茶和茶梅的生长指标影响的双因素方差分析Table 2 Two-way ANOVA of the effects of nitrogen deposition rate and addition frequency on the growth indicators of Camellia japonica(NaiDong) and Camellia sasanqua
2.2 耐冬山茶和茶梅的生物量对不同氮沉降的响应
耐冬山茶幼苗的生物量不受氮沉降和添加频率及两者交互作用处理的影响,而茶梅幼苗的叶生物量(MLB)和总生物量(MTB)在不同添加频率的作用下差异显著,其他因素对茶梅生物量的影响均不显著(表3)。在轻度氮沉降和重度氮沉降处理下茶梅的MLB、MTB都随着添加频率的提高而升高(图2)。

图2 氮沉降量和氮沉降添加频率的改变对植株生物量的影响Fig. 2 Effects of nitrogen deposition rate and nitrogen deposition frequency on biomass parameters of plant
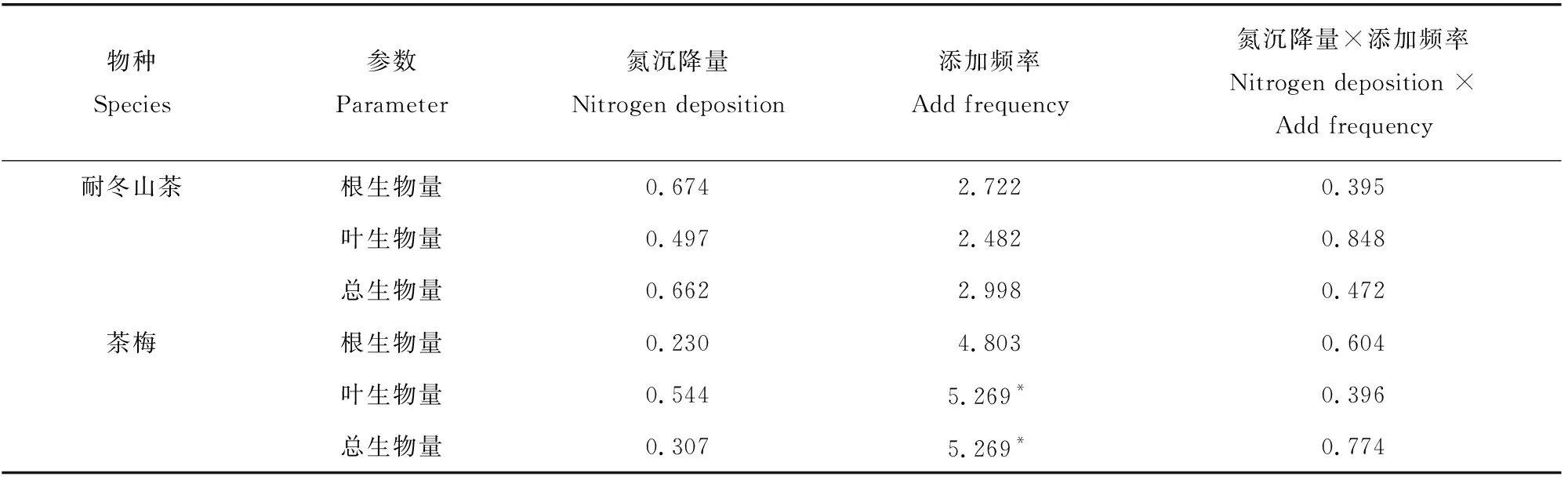
表3 氮沉降量和添加频率对耐冬山茶和茶梅的生物量影响的双因素方差分析Table 3 Two-way ANOVA of the effects of nitrogen deposition rate and addition frequency on biomass of Camellia japonica(NaiDong) and Camellia sasanqua
2.3 耐冬山茶和茶梅的生物量比对不同氮沉降的响应
由表4可知,添加频率极显著地影响了茶梅的根生物量比(RMR),氮沉降量对茶梅幼苗的根冠比(RS)影响显著。添加频率对耐冬山茶幼苗的RMR也有显著影响。在相同的添加频率下,茶梅幼苗在轻度氮沉降量处理组下的RS均为最低值;在相同氮沉降处理下,低添加频率茶梅幼苗的RMR均达到最高值。耐冬山茶在各个氮沉降量处理下,RMR均在中添加频率时达到最大值(图3),分别为0.61、0.46、0.77。
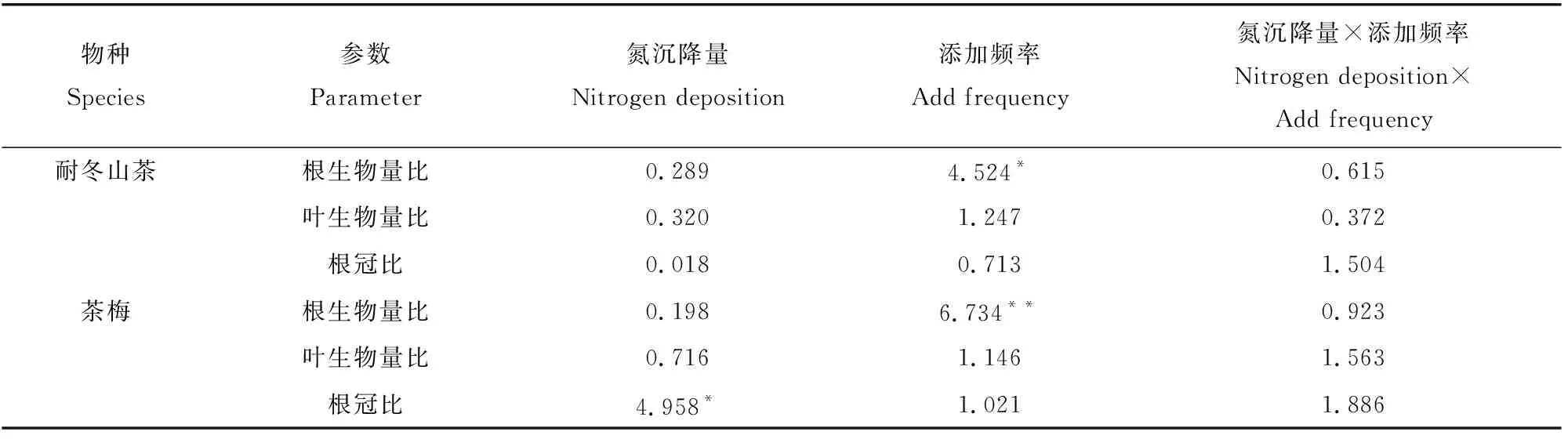
表4 氮沉降量和添加频率对耐冬山茶和茶梅的生物量分配影响的双因素方差分析Table 4 Two-way ANOVA of the effects of nitrogen deposition rate and addition frequency on biomass distribution of Camellia japonica(NaiDong) and Camellia sasanqua

图3 氮沉降量和氮沉降添加频率的改变对植株生物量分配的影响Fig. 3 Effects of nitrogen deposition rate and nitrogen deposition frequency on biomass distribution of plant
2.4 耐冬山茶和茶梅的气体交换参数对不同氮沉降的响应
由表5可知,氮沉降量除了对茶梅幼苗的净光合速率(Pn)影响不显著、对蒸腾速率(E)影响显著外,对茶梅幼苗的细胞间隙CO2浓度(Ci)、气孔导度(Gs)和气孔限制值(LS)及耐冬山茶幼苗的气体交换参数的影响极其显著;氮沉降量和添加频率互作对茶梅Ci和LS有显著影响,对山茶影响不显著;添加频率对两种植物的气体交换参数均无显著影响(表5)。由图4可知,在同一添加频率下,耐冬山茶Pn随着氮沉降的增加先上升后下降。高添加频率和中添加频率下,随着氮沉降的增加,Ci先下降后上升,低添加频率处理下Ci呈下降趋势,相较于对照组和轻度氮沉降量处理组分别下降了51.65%和22.62%。E在高添加频率和低添加频率时,随着氮沉降的增加而呈下降趋势,中添加频率处理下,随着氮沉降的增加先上升后下降;在同一添加频率下,对照组和轻度氮沉降组基本相似,但重度氮沉降组相较于轻度氮沉降组分别下降了15.72%、18.22%、16.31%。耐冬山茶Gs在相同添加频率处理下,随着氮沉降的增加而降低;耐冬山茶LS在高添加频率和中添加频率的处理下,随着氮沉降的增加先上升后下降,低添加频率处理下随着氮沉降的增加而上升。茶梅在高添加频率处理下的Ci随着氮沉降的增加呈下降趋势,相较于对照组,轻度氮沉降和重度氮沉降分别下降了44.34%和52.76%,而在中、低添加频率下随着氮沉降的增加Ci先下降后上升。高添加频率下茶梅E和Gs随着氮沉降的增加呈下降趋势,中、低添加频率下降趋势明显变缓。在高添加频率的处理下,随着氮沉降的增加,茶梅LS呈上升趋势,而在中添加频率和高添加频率处理下先上升后下降。
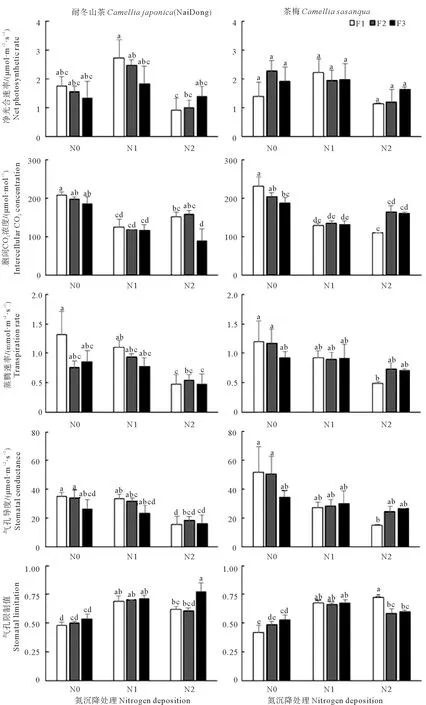
图4 氮沉降量和氮沉降添加频率的改变对植株气体交换参数的影响Fig. 4 Effects of nitrogen deposition rate and nitrogen deposition frequency on gas exchange parameters of plant

表5 氮沉降量和添加频率对耐冬山茶和茶梅的气体交换参数影响的双因素方差分析Table 5 Two-way ANOVA of the effects of nitrogen deposition rate and addition frequency ongas exchange parameters of Camellia japonica(NaiDong) and Camellia sasanqua
2.5 耐冬山茶和茶梅的叶绿素荧光对不同氮沉降的响应
初始荧光F0是光系统Ⅱ(PSII)光化学反应中心处于完全开放时的荧光产量,F0高低反映了PSII反应中心的活性和植物体色素含量的多少。PSⅡ反应中心的潜在活性(Fv/F0)反映了PSⅡ光化学效率。根据表6,耐冬山茶F0和Fv/F0受氮沉降量和添加频率的交互作用影响分别为显著和极显著;氮沉降量仅对茶梅的F0有显著影响。由图5知,耐冬山茶F0在轻度氮沉降和高添加频率的条件下高于其他处理组;Fv/Fm在非胁迫环境下变化极小,而且不受物种和生长条件影响,试验中各处理该值接近于0.8~0.84,说明植株叶片为健康叶。轻度氮沉降中、低添加频率的条件下的耐冬山茶Fv/F0显著高于同氮沉降量的高添加频率组,这与上述分析的Fv/F0受添加频率及二者交互作用的影响显著一致。茶梅幼苗F0在中添加频率处理下,重度氮沉降量处理组与对照组和轻度氮沉降处理组相比分别下降25.84%和24.24%。

表6 氮沉降量和添加频率对耐冬山茶和茶梅的叶绿素荧光影响的双因素方差分析Table 6 Two-way ANOVA of the effects of nitrogen deposition rate and addition frequencyon chlorophyll fluorescence of Camellia japonica(NaiDong) and Camellia sasanqua
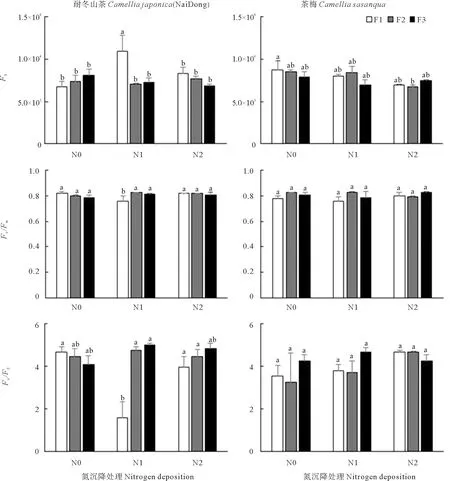
图5 氮沉降量和氮沉降添加频率的改变对植株叶绿素荧光的影响Fig. 5 Effects of nitrogen deposition rate and nitrogen deposition frequency on chlorophyll fluorescence parameters of plant
3 讨论
氮元素作为植物生长发育过程中必不可少的元素,对植物具有重要意义,影响植物的生长与发育。氮元素的添加频率同样影响植株的生长与分布,添加频率的不同对植株产生的影响不同。
本研究中氮沉降量和添加频率的交互作用对耐冬山茶幼苗和茶梅幼苗大部分指标的影响并不显著,测定的大部分参数仅仅受氮沉降或添加频率的单因素影响。氮沉降量影响了耐冬山茶幼苗和茶梅幼苗的光合作用,氮沉降和氮沉降添加频率的交互作用没有对耐冬山茶幼苗的气体交换参数产生显著影响,对茶梅幼苗的影响集中在Ci和LS,因此可以推断,不同的添加频率能够改变植株对氮沉降的响应策略[12]。
在光合作用中氮元素的地位极其重要,在蛋白质和叶绿素的合成中不可或缺,因此氮沉降对植物进行光合起到了积极的促进作用,使有机物的积累增加[11, 13]。Villar-Salvador等[14]、Fang等[15]的研究结果显示,高于正常水平的氮沉降会存在促进作用,使有机物积累;另一种观点则指出,过量的氮元素会不利于生物量的积累,使生产力下降[9, 16-17]。总体来看,耐冬山茶幼苗的基径在N0(0 g·m-2·a-1)、N1(6 g·m-2·a-1)水平下显著高于N2(12 g·m-2·a-1),而茶梅幼苗的冠面积在N0水平下显著高于N1、N2水平。因此,当氮沉降达到N1水平时,茶梅幼苗的生长受到了限制,当氮沉降达到N2水平时,对耐冬山茶幼苗的生长有抑制作用,这说明了氮沉降对耐冬山茶和茶梅的促进作用存在阈值,且耐冬山茶的阈值要高于茶梅。这种阈值效应可能需要更长时间才能反映到两物种的生物量中。当氮沉降较低时,植物的生长发育会受到促进作用,一旦氮沉降的速率升高超过阈值,其促进作用就会下降,这一结果与前人对元宝枫(Acertruncatum)的研究相一致[16-17]。耐冬山茶幼苗和茶梅的株高不受氮沉降增加的影响,这与之前的研究结果相反[6, 9, 16],这可能与叶的其他性状的变化相关,比如叶片的Pn、叶绿素含量等。试验结果表明耐冬山茶幼苗和茶梅幼苗的生长指标受氮沉降加速的影响不大,与麻栎(Quercusacutissima)、蒙古栎(Quercusmongolica)等植物的研究结果相一致[6, 9],但却与生菜(Lactucasativa)等植物的研究结论相悖[18],这种差异可能是由于不同植物对氮的需求量、敏感程度不同和生长速度迥异造成的。
添加频率影响了茶梅幼苗的生物量,但耐冬山茶的生物量却不受添加频率的影响。较高的添加频率能够显著地增加植物的生物量[2],茶梅的试验结果也证实了这一观点。Fransen等人[19]通过对多年生植物绒毛草和甘松茅的研究发现,添加频率的改变虽然能使植物在短期时间内通过大量根系的增生使植物获取大量的养分,促进植物地上部的生长,但从长期的生长来看这种优势很快会消失。由此可以看出,相较于耐冬山茶,茶梅的生物量对添加频率的响应更为敏感,推测可能是因为耐冬山茶的根系相较于茶梅更为发达,在低营养物质的环境中,不需要通过增加根系来吸取更多的养分;而茶梅的根系在短时间内通过增生获取大量养分保证生长,从而使茶梅的根生物量比在短时间内急速上升。
在氮沉降初期耐冬山茶和茶梅对短时间的氮沉降比较敏感,并且不同的植物对氮沉降呈现出不同的响应可能是因为不同的植物对于氮的敏感度存在差异。Farquhar和Sharkey[20]认为:如果植株表现出Pn下降的同时E和Ci也一起下降,则为气孔因素的原因;如果是非气孔因素为主要原因,则Pn会随着Ci的上升而下降,也就是说因为叶肉细胞光合活性下降而导致了Pn的下降。本研究中耐冬山茶幼苗的Pn升高时,Ci下降;而耐冬山茶幼苗的Pn降低时,Ci上升,说明了耐冬山茶幼苗的Pn受非气孔因素限制,主要是由于叶肉细胞光合活性的下降造成的。氮沉降除了对茶梅植株的Pn影响不显著之外,对两物种的其他光合参数均影响显著,这一结果与元宝枫的研究结果一致[16-17]。整体上耐冬山茶随着氮沉降的增加,Pn、E先升后降,Ci先降后升,Gs下降,LS上升;而茶梅随着氮沉降的增加,Ci先降后升,E、Gs下降,Ls先升后降。氮沉降添加频率发生改变,对山茶和茶梅的生长与气体交换并无显著影响,由此推测:一是耐冬山茶幼苗和茶梅幼苗并不需要很多的氮元素,原有的氮元素含量就已经足够,更高的氮沉降并不会使其做出响应;二是氮流失导致氮沉降加速并未形成胁迫;三是耐冬山茶幼苗和茶梅幼苗的生长发育缓慢,短期处理不会产生影响。叶绿素荧光参数是研究植物光合作用的探针,能反映出植物进行光合作用过程中的变化[19-21]。如果PSⅡ反应中心遭受破坏则F0会表现出增加。在本试验中,耐冬山茶F0在氮沉降条件下,随着添加频率的增加而上升,说明较高的添加频率条件下PSⅡ反应中心遭受影响,但并没有造成不可逆性失活或破坏。耐冬山茶幼苗叶片的Fv/F0在中、高氮沉降量下随着添加频率的降低显著提高,说明随着添加频率的降低,叶片的PSⅡ反应中心的潜在活性提高。耐冬山茶及茶梅的Fv/Fm未受添加频率的影响,数值均维持在0.8~0.84之间,说明了添加频率对两种植物的叶绿素最大量子产量无明显影响。综上所述,耐冬山茶和茶梅在受到不同氮沉降胁迫时,会通过改变自身的生长性状和生理活动等抵御或适应逆境,因此耐冬山茶和茶梅对氮沉降具有一定耐受性。氮沉降的促进作用可能存在阈值,且耐冬山茶的阈值可能明显高于茶梅,过高的氮沉降会对耐冬山茶和茶梅产生抑制作用。在当前氮沉降日益严重的环境下,耐冬山茶和茶梅适合作为园林绿化树种继续使用,在氮沉降量较低的地区可以通过增加氮肥的方式促进植株的生长,较高的降雨频率有助于缓解高氮沉降的消极影响。
猜你喜欢
今日农业(2021年10期)2021-11-27 09:45:24
今日农业(2020年19期)2020-12-14 14:16:52
文苑(2020年6期)2020-06-22 08:41:50
大观(书画家)(2018年6期)2018-07-08 00:43:20
民族音乐(2016年1期)2016-08-28 20:02:52
中国科技信息(2015年2期)2015-11-16 08:18:32
中国火炬(2015年12期)2015-07-31 17:38:35
小溪流(画刊)(2014年6期)2014-08-11 19:25:34
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
中国火炬(2013年3期)2013-07-24 14:15:06
