
Hydrogen sulfide enhances adult neurogenesis in a mouse model of Parkinson’s disease
2021-12-09 14:12MinWangJuanJuanTangLinXiaoWangJunYuLiZhangChenQiao
中国神经再生研究(英文版) 2021年7期
Min Wang , Juan-Juan Tang , Lin-Xiao Wang, Jun Yu, Li Zhang , Chen Qiao
Abstract Hydrogen sulfide (H2S) is regarded to be a protectant against diseases of the central nervous system and cardiovascular system. However, the mechanism by which H2S elicits neuroprotective effects in the progression of Parkinson’s disease (PD) remains unclear. To investigate the role of H2S in delaying the pathological process of PD, we used the most common sodium hydrosulfide (NaHS) as an H2S donor and established a mouse model of PD induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine/probenecid (MPTP/p) in the present study. Our results show that H2S reduced neuronal loss during the progression of PD. Notably, we found that H2S exhibited protective effects on dopaminergic neurons. Excitingly, H2S also increased the proliferation of neural stem cells in the subventricular zone. Next, we evaluated whether the neuroprotective effects of H2S on dopaminergic neurons in PD are dependent on adult nerve regeneration by treating primary adult neural stem cells cultured ex vivo with 1-methyl-4-phenylpyridine. Our results show that H2S could prevent nerve injury induced by 1-methyl-4-phenylpyridine, promote the growth of neurospheres, and promote neurogenesis by regulating Akt/glycogen synthase kinase-3β/β-catenin pathways in adult neural stem cells. These findings confirm that H2S can increase neurogenesis in an adult mouse model of PD by regulating the Akt/glycogen synthase kinase-3β/β-catenin signaling pathway. This study was approved by the Animal Care and Use Committee of Nanjing Medical University, China (IACUC Approval No. 1601153-3).
Key Words: factor; hydrogen sulfide; medical gas; neural stem cells; neurogenesis; neuroprotection; Parkinson’s disease; pathways
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disease after Alzheimer’s disease, which seriously affects the quality of life of older people (Aarsland et al., 2017; Baiano et al., 2020). Characteristics of PD include the progressive degeneration of dopaminergic (DA) neurons in the substantia nigra pars compacta (SNc) of the midbrain,which leads to dyskinesia such as muscle rigidity, bradykinesia,and tremors in PD patients (Aarsland et al., 2017; Benzagmout et al., 2019; Yakhine-Diop et al., 2019). At present, the most commonly accepted theory is that oxidative stress (Trist et al.,2019), mitochondrial dysfunction (Elfawy and Das, 2019), and accumulation of α-synuclein (Tan et al., 2020) are the primary factors inducing the loss of DA neurons. However, the precise mechanistic relationships that contribute to PD and the efficacy of therapeutics used to treat PD remain obscure. Thus, there is an urgent need to develop medications that can effectively relieve the symptoms of PD and slow the loss of DA neurons.
Hydrogen sulfide (H2S) can act in the body as a gasotransmitter, a gaseous signaling molecule that can transmit signals and induce physiological responses in various tissues, and exists widely in mammals (Cao et al.,2018; Pushchina et al., 2020). In recent decades, H2S has been connected with both physiological and pathological roles in multiple systems, such as the cardiovascular (Zhang et al., 2018), respiratory (Guan et al., 2020), and central nervous systems (Cao et al., 2018; Ma et al., 2019). Increasing evidence shows that H2S exerts anti-oxidant (Ali et al., 2019),anti-inflammatory (Wang et al., 2017), and anti-apoptotic effects in the brain (Wang et al., 2017, 2019). In the midbrain,the major enzymes involved in endogenous H2S biosynthesis are cystathionine β-synthase and cystathionine γ-lyase (Kimura et al., 2015). As the midbrain is the dominating brain region affected in the pathogenesis of PD, H2S has been suggested to be beneficial in attenuating PD-like neural damage (Cakmak,2016; Yuan et al., 2018). Nevertheless, the relationship between protective mechanisms of H2S and the pathogenesis of PD remains largely elusive.
Adult neurogenesis is one method to combat the loss of DA neurons in the adult midbrain, which may prevent cell death and relieve dyskinesia in PD (Farzanehfar, 2018; Hain et al., 2018). The subgranular zone and subventricular zone(SVZ) are the two main neurogenic regions in the adult brain (Westerlund et al., 2005). Since the discovery of these important centers of regeneration, adult neurogenesis in the SVZ has been an area of active research (Fan et al., 2016;Dillen et al., 2020). Emerging evidence indicates that fresh DA neurons generated by neural stem cells (NSCs) in the SVZ could significantly enhance the functional performance of patients, suggesting that neurogenesis may be a feasible therapeutic strategy in adults (Le Grand et al., 2015; Fan et al., 2016; Farzanehfar, 2018). Previous studies have focused on DA neurons and how to delay neuronal degeneration by treatment with H2S (Wang et al., 2015; Yin et al., 2017).However, it remains unclear whether H2S therapy could induce neurogenesis in adults to alleviate the progression of PD.
Herein, we utilized sodium hydrosulfide (NaHS) as a donor of H2S (Cao et al., 2018; He et al., 2019), and further investigated the role of H2S in PD using a classical 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) model.
Materials and Methods
Animals
All studies were approved by the Animal Care and Use Committee of Nanjing Medical University, China (IACUC Approval No. 1601153-3) on August 6, 2016. Thirty-six specific-pathogen-free adult male C57BL/6J mice aged 12–16 weeks and weighing 26–32 g, were applied in thein vivostudy.Another 36 specific-pathogen-free male C57BL/6J mice aged 2–3 months were used for NSC isolation in anin vitrostudy. All mice were purchased from the Laboratory Animal Center of Nanjing Medical University (license No. SCXK (Su) 2013-0017).The mice were housed with free access to food and water in a room with an ambient temperature of 22 ± 2°C and a 12:12-hour light/dark cycle.
Animal model preparation
A total of 36 mice were randomly divided into three groups:saline, MPTP, and MPTP + NaHS (n= 12 mice per group). One hour after intraperitoneal injection of probenecid (250 mg/kg;Jinan Times Pharmacology, Jinan, China), which was dissolved in dimethyl sulfoxide 30 minutes before injection, MPTP (20 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) was subcutaneously injected daily for 5 days to establish the PD model. In the NaHS group, mice were intraperitoneally injected with NaHS (5.6 mg/kg; Sigma-Aldrich) for 10 days (from days 4 to 8), 30 minutes before MPTP injection (Figure 1A). The saline group was injected with the same volume (about 0.4 mL) of normal saline for 10 days. At 3.5 days after the last administration of MPTP, mice were sacrificed by cervical dislocation or 5% chloral hydrate.
Nissl staining
Brain tissues were immobilized in 4% paraformaldehyde,which was prepared in 0.1 M phosphate buffer (pH 7.4) at 4°C.Midbrain tissue [according to the Atlas of Paxinos and Franklin(2013), from approximately –2.5 to –3.88 mm] was prepared into 30-µm-thick slices using a Leica cryostat (CM1860;Leica, Germany). Slices were stained with cresyl violet acetate, photographed, and counted by MicroBrightField Stereo Investigator software (MBF Bioscience, Williston, ND,USA). All histological section count data were obtained by three observers in a randomized double-blind controlled experiment.
Bromodeoxyuridine labeling
On the last day of the 5-day MPTP/probenecid cycle, mice were intraperitoneally injected with bromodeoxyuridine[BrdU; Sigma-Aldrich; 50 mg/kg four times with an interval of 2 hours (Sairanen et al., 2005)]. After the last BrdU treatment, mice were transcardially perfused with 0.1 M phosphate-buffered saline (PBS) for 5 minutes, followed by 4%paraformaldehyde. Four weeks after injection with BrdU, mice were sacrificed and perfused, and brain tissue was frozen at–70°C for subsequent calculation of the differentiation and survival of BrdU-labeled cells.
Immunofluorescence staining
The immunostaining method utilized has been described in previous literature (Qiao et al., 2017). Mice were infused with 4% paraformaldehyde after model preparation. Next,brain tissues were post-fixed in 4% paraformaldehyde at 4°C overnight, followed by transfer into 15% sucrose in PBS overnight, then 30% sucrose overnight, until the brain sank to the bottom of the tube. Brain tissue was sliced into 20-µmthick slices using a cryostat. Brain slices were washed with PBS for 10 minutes, blocked with 5% bovine serum albumin in PBS for 1 hour, and then incubated with the appropriate primary antibody [anti-tyrosine hydroxylase (TH), a ratelimiting enzyme in the biosynthesis of dopamine, Cat# T1299,Sigma-Aldrich; anti-BrdU, a new-cell marker, Cat# MAB3510,Millipore, Boston, MA, USA; anti-Ki67, a proliferating cell marker, Cat# 610968, Becton Dickinson, Franklin Lakes, NJ,USA) overnight at 4°C. The second day, slices were incubated with the corresponding fluorescent secondary antibody [goat anti-mouse IgG-fluorescein isothiocyanate (Cat# ab150113,Abcam, Cambridge, UK) and goat anti-rabbit IgG-fluorescein isothiocyanate (Cat# ab150077) for 1 hour. Nuclei were stained with 4′,6-diamidino-2-phenylindole dye.
Image analysis
Images acquired under a confocal light microscope(Axiovert LSM510; Carl Zeiss Co., Weimar, Germany) were quantitatively analyzed for immunostaining signals using Optical fractionation and MicroBrightField Stereo Investigator software. Total numbers of Nissl+neurons and TH+neurons in the SNc, as well as Ki67+cells and BrdU+cells in the SVZ,were calculated from six different samples in each group.Before placing the coverslips, a fluorescent mounting medium was applied and allowed to dry in the dark. Specimens were examined with MicroBrightField Stereo Investigator software for counting and photography.
Adult NSC culture
Mouse adult NSCs (ANSCs) were cultured according to an established primary culture system (Fan et al., 2016). In brief, coronal section microanatomy was performed on the brains of male C57BL/6J mice aged 2–3 months to obtain the periventricular rostral region. Tissues were gently chopped and incubated with papain hydrolysate at 37°C for 30 minutes. The supernatant was carefully discarded after centrifugation, and cells were re-suspended in a serum-free medium consisting of Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12(1:1) (Gibco, Grand Island, NY, USA), 20 ng/mL epidermal growth factor (Peprotech, Rocky Hill, CT, USA), 2% B27 (Gibco),and 20 ng/mL basic fibroblast growth factor (Peprotech). The main cell culture technique used in this research was the neurosphere measure (Kong et al., 2008). Cell viability was evaluated by trypan blue exclusion (0.4%; Sigma-Aldrich), and exceeded 85%. In uncoated 24-well culture plates, the primary tissue was seeded at a rate of 10,000 cells per well. After 7 days of culture, the size of neurospheres and total number formed in each well were calculated. Self-renewal was also carried out by separating large numbers of neurospheres and re-culturing the cells at a constant density of 10,000 cells per well. After 7 days of culturein vitro, the number and size of second- or third-level spheres was evaluated. Under a confocal light microscope, neurospheres with less than a minimum diameter of 40 µm were excluded. Diameters of randomly selected neurospheres were measured by Image-Pro Plus software (Media Cybernetics, Inc., Bethesda, MD, USA).
Cell proliferation detection
Proliferation was measured using a BrdU-incorporation method (Fan et al., 2016). In brief, we collected neurospheres in control, 1-methyl-4-phenylpyridiniumion (MPP+), and MPP++ NaHS groups, and gently mechanically separated them.Separated cells were cultured for 48 hours in 24-well plates pretreated with poly-L-ornithine (10 µg/mL) and laminin (5µg/mL). Neurospheres were cultured with 10 µM BrdU for 60 minutes and then fixed. After washing, cells were treated with 2 M HCl at 37°C for 30 minutes to denature the DNA. After washing, cells were blocked with 5% bovine serum albumin for 40 minutes, incubated at 37°C for 1 hour with primary mouse anti-BrdU antibody (1:5000; Millipore), and then stained with anti-mouse antibody conjugated to fluorescein isothiocyanate(1:1000; Invitrogen, Carlsbad, CA, USA) at 37°C for 1 hour.Nuclei were stained by 4′,6-diamidino-2-phenylindole and mounted with anti-fade medium. Under a confocal light microscope, images were acquired of 10 randomly selected visual fields; fluorescence intensity of images was calculated with Image-Pro Plus software (Media Cybernetics, Inc.).
Western blot assay
Proteins within neurospheres were extracted with homogenate buffer. An equivalent amount of protein (50 µg)was transferred to a polyvinylidene difluoride membrane by sodium dodecyl sulfate-polyacrylamide gel electrophoresis.After blocking membranes with 5% skimmed milk in PBS, they were incubated with primary antibodies including rabbit anti-proliferating cell nuclear antigen [PCNA; 1:1000; Cat# 13110;Cell Signaling Technology (CST), Boston, MA, USA], rabbit anti-Akt (1:1000; Cat# 2920; CST), rabbit anti-pAkt (1:1000; Cat#12694; CST), rabbit anti-H3 (1:500; CST), rabbit anti-β-catenin(1:1000; Cat# 8480; CST), rabbit anti-p-β-catenin (1:1000; Cat#2009; CST), mouse anti-glycogen synthase kinase-3β (GSK-3β; 1:1000; Cat# 21301; SAB, Nanjing, China), and mouse anti-phosphorylated GSK-3β (p-GSK-3β; 1:1000; Cat# 11301;SAB) overnight at 4°C. According to the manufa cturer’s instructions, immunoreactive proteins were examined using horseradish peroxidase-labeled secondary antibodies and ECL kits (Amersham Biosciences, Piscataway, NJ, USA). Finally, we scanned and analyzed membranes by using an Omega 16ic Chemiluminescence Imaging System (Ultra-Lum, Claremont,CA, USA).
Statistical analysis
All the data are shown as mean ± standard error of the mean(SEM). One-way analysis of variance followed by Tukey’spost hoctest was used to evaluate the statistical significance of results using Graphpad Software 5.0 (GraphPad Software,San Diego, CA, USA).P-values less than 0.05 were considered statistically significant.
Results
H2S rescues the loss of DA neurons in an MPTP mouse model of PD
To determine the role of H2S in an MPTP mouse model of PD, we used Nissl staining to evaluate neuronal injury. As demonstrated inFigure 1B, compared with the saline group,MPTP treatment decreased the total number of neurons in the SNc by 39% (F(2,15)= 106.2,n= 6,P< 0.0001), but treatment with 5.6 mg/kg NaHS led to a loss of just 9% of neurons (F(2,15)= 106.2,n= 6,P=0.0075;Figure 1BandC).Furthermore, we observed damage to DA neurons using TH immunostaining. In this study, we found that MPTP observably reduced the number of TH-positive cells by 43% in the SNc(F(2,15)= 81.51,n= 6,P< 0.0001;Figure 2AandB). Similar to the results of Nissl staining, NaHS treatment clearly decreased the loss of TH-positive cells caused by MPTP, leading to only a 16% loss relative to the Saline group (F(2,15)= 81.51,N= 6,P= 0.0008;Figure 2B). These results indicate that H2S has protective effects in the MPTP mouse model of PD, especially with regard to the loss of DA neurons.
H2S attenuates inhibition of SVZ neurogenesis in the MPTP mouse model
Neurogenesis is one potential mechanism to reverse DA cell loss in the SNc of MPTP-induced PD (Lindvall and Kokaia, 2009).To evaluate the protective effect of H2S on DA neurons, the extent of neurogenesis was evaluated. Using immunostaining for Ki67, an endogenous label of cellular proliferation(Fan et al., 2016; Farzanehfar, 2018), we discovered that Ki67-positive cells were enriched by 50% (F(2,15)=121.0,n= 6,P< 0.0001) in the SVZ following treatment with NaHS. However, Ki67-positive cells were decreased by 58%after MPTP treatment alone (F(2,15)= 114.4,n= 6,P< 0.0001;Figure 3AandB). There was no significant change in Ki67 expression with NaHS treatment alone (Additional Figure 1). A BrdU immunoassay was used to further illustrate the role of H2S in increasing cell proliferation (Fan et al., 2016;Farzanehfar, 2018). We noticed a meaningful reduction of BrdU-positive cells in the SVZ following treatment with MPTP(F(2,15)= 62.89,n= 6,P< 0.0001). However, consistent with the results of Ki67 staining, H2S treatment markedly increased the number of BrdU-positive cells in MPTP mice (F(2,15)= 62.89,n=6,P< 0.0001;Figure 3CandD). Thus, we found that H2S could promote neurogenesis and restore the loss of cell proliferation in the SVZ induced by MPTP.
H2S promotes the proliferation of ANSCs injured by MPP+ in vitro
To further explore the role of H2S in promoting cell proliferation, we cultured adult ANSCsin vitro. As shown inFigure 4A, the number and diameter of neurospheres isolated from the SVZ brain region of WT mice were calculated.We observed no significance differences in the number of neurospheres between control and MPP+-treated groups. In addition, NaHS treatment could slightly increase the number of neurospheres, but had no statistically significant effect (F(2,9)=1.386,P> 0.05;Figure 4B). Nevertheless, MPP+inhibited the growth of neurospheres and reduced their diameter by 30%compared with the control group (F(2,9)= 13.43,P= 0.0018).Unexpectedly, NaHS treatment alleviated the negative effects of MPP+on neurospheres and restored the diameter to 91%of untreated groups (F(2,9)= 13.43,P= 0.3636;Figure 4C).
Furthermore, a BrdU proliferation assay was used to measure growth of the neurospheres (Fan et al., 2016; Farzanehfar,2018). BrdU-positive cells were decreased by 52% in MPP+-treated ANSCs (F(2,9)= 25.61,P= 0.0003). However, NaHS treatment restored proliferation (F(2,9)= 25.61,P= 0.0007),leading to only a 6% loss in positive cells relative to the control group (F(2,9)= 25.61,P= 0.7154;Figure 5AandB).In conclusion, H2S had no effect on the number of cultured ANSCs, but dis increase cell proliferation to promote the growth of ANSCs.
H2S enhances ANSC proliferation via the Akt/GSK-3β/β-catenin pathway
To assess the role of H2S in proliferation of cultured ANSCs, protein was extracted from neurospheres and the concentration of PCNA was measured in the nucleus. PCNA was downregulated in the nuclei of MPP+-treated ANSCs,however treatment with NaHS reversed this effect, leading to an increase in PCNA expression (F(2,9)= 15.75,P= 0.0191;Figure 6AandB). Consistent with previous experiments,these observations corroborate the role of H2S in enhancing neurogenesis in the MPTP model of PD.
To explore the mechanisms by which H2S affects neurogenesis,western blotting was performed to identify differentially regulated signaling molecules. As PCNA is a target of Akt signaling, which is frequently activated in cell proliferation(Bertapelle et al., 2017), we hypothesized that the neuroprotective roles of NaHS may be promoted by Akt signaling. As shown inFigure 6AandC, MPP+remarkably decreased expression of phosphorylated Akt (p-Akt) in NSCs by 59% (F(2,9)= 43.84,P< 0.0001), however, this effect was attenuated by treatment with NaHS (F(2,9)= 43.84,P= 0.0002;Figure 6C). This result supports the potential role of H2S in promoting neurogenesis through the Akt pathway.
According to the latest studies, GSK-3β is involved in the canonical Wnt signaling pathway and controls the level of β-catenin phosphorylation, thereby enhancing the selfrenewal and proliferation potential of NSCs (Fidaleo et al.,2017; Horgusluoglu et al., 2017). Thus, our next step was to measure the contents of phosphorylated GSK-3β and β-catenin to evaluate whether H2S affects this pathway. In extracts of ANSCs, pGSK-3β was significantly increased after stimulation of MPP+ (F(2,9)= 77.28,P< 0.0001;Figure 6DandE), but the level of p-β-catenin was markedly decreased (F(2,9)= 34.63,P< 0.0001;Figure 6F). However, after treatment with H2S,expression of pGSK-3β and p-β-catenin was reversed, with a decrease in pGSK-3β (F(2,9)= 77.28,P< 0.0001;Figure 6E) and an increase in p-β-catenin (F(2,9)= 34.63,P= 0.0004;Figure 6F) expression compared with their total proteins (Figure 6D).These results support for the idea that H2S could influence the Akt/GSK-3β/β-catenin cascade to promote adult neurogenesis in PD.
Discussion
The SVZ is the main proliferative region of the adult brain.Accumulating evidence indicates that DA neurons produced by NSCs in the SVZ can measurably improve PD symptoms(Horgusluoglu et al., 2017). H2S is a gas that has been shown to have neuroprotective effects in PD. Nevertheless, the precise mechanisms by which H2S acts against PD remain unclear. Treatment with NaHS instantaneously generates H2S via pH-dependent salt dissociationin vivoandin vitro. In the present study, we used NaHS to produce H2S and showed that H2S could not only delay the loss of DA neurons in the SNc, but also further promoted adult neurogenesis in the SVZ of PD model animals. Further experiments suggested that H2S remarkably increased the proliferation and growth of ANSCs culturedin vitro. Excitingly, we found that H2S may regulate the Akt/GSK-3β/β-catenin cascade to enhance the growth of ANSCs. Together, these findings demonstrate a new neuroprotective role for H2S in promoting adult neurogenesis in PD.
H2S is an abundant gas in nature that plays a vital role as a signaling element in the central nervous system. The benefits of H2S in PD have recently been confirmed by accumulating reports from other labs (Cao et al., 2018; Shefa et al., 2018).Herein, we identified the molecular function of H2S in a PD model. Our results demonstrate that H2S rescued the loss of DA neurons following MPTP stimulation. Previous studies have demonstrated anti-apoptotic, anti-oxidative, and antiinflammatory roles of H2S in PD models (Cao et al., 2018;Sarukhani et al., 2018). Nevertheless, the specific mechanism underlying the neuroprotective effects of H2S on PD has remained largely elusive.
Emerging evidence demonstrates that adult neurogenesis is beneficial for the recovery of neurons in the pathological process of PD. In the mammalian brain, the two major neurogenic zones are the SVZ of the lateral ventricles and subgranular zone of the dentate gyrus in the hippocampus(Horgusluoglu et al., 2017). Adult neurogenesis in the SVZ is mostly associated with migration of neuroblasts along the rostral migratory stream and generation of neurons in the striatum (Fidaleo et al., 2017). Expression of Ki67, a nuclear protein expressed primarily in proliferating cells (Fan et al.,2016), was fortified in the SVZ by H2S in the presence of MPTP stimulation. This result suggests that the protective effect of H2S on DA neurons may depend on promotion of nerve cell proliferation. We further found that after treatment with H2S,BrdU staining significantly increased, indicating an enhanced number of proliferating cells in the SVZ. To confirm the role of H2S in adult neurogenesis, we cultured mouse ANSCs from the SVZ under MPP+stimulation. Following pre-treatment with H2S, we noted that MPP+-induced inhibition of neurosphere growth was decreased, but there was no effect on the number of neurospheres. However, as expected, BrdU-labeled proliferating cells were significantly increased in cultured neurospheres following treatment with H2S. Accordingly, we clarified that H2S regulates adult neurogenesis to postpone DA neuronal degeneration in models of PD.
The Akt/GSK-3β/β-catenin cascade has recently emerged as a pivotal step in the proliferation of NSCs in the central nervous system (Fidaleo et al., 2017). Therefore, we hypothesized that H2S may control activity of the Akt/GSK-3β/β-catenin cascade to resist damage induced by MPP+in ANSCs. Here, we found that the Akt pathway was activated and p-GSK-3β expression levels were decreased after H2S treatment. Notably, a recent study indicated that GSK-3β can be inactivated by p-Akt, and p-GSK-3β stabilizes β-catenin, which then migrates into the nucleus to mediate gene transcription enabling neurogenesis.In our study, we observed increased p-β-catenin levels in the group pre-treated with H2S compared with MPP+treatment alone. Consistently, PCNA expression was highly upregulated along with p-β-catenin expression levels. Our findings provide evidence that H2S exerts a latent role in facilitating adult SVZ neurogenesis by activating the Akt/GSK-3β/β-catenin cascade to increase related gene transcription.
In conclusion, we demonstrated that H2S exerts neuroprotective roles in a pathological mouse model of PD. Furthermore,H2S promoted adult neurogenesis in areas of the SVZ that had been damaged by MPTP or MPP+. Most notably, our study showed that the benefits of H2S in accelerating adult neurogenesis may be mediated by Akt/GSK-3β/β-catenin signaling activation. Further work is required to investigate whether proliferating ANSCs could be differentiated into DA neurons capable of making functional synaptic connections with neurons in a PD model or α-synuclein overexpression model. These findings will further increase our recognition of the potential of H2S as a new therapy for PD.
Author contributions:Conceptualization: CQ, LZ; methodology: MW, JJT;investigation: MW, JJT, LXW, JY, CQ, MW; writing original draft: CQ, MW;manuscript editing: JJT, LZ; fundraising: LXW, JY, CQ; supervision: CQ;project administration: CQ, LZ. All authors contributed to this study and approved the final manuscript.
Conflicts of interest:The authors declare no conflict of interest.

Figure 1|Effects of H2S on MPTP-induced PD mouse model.

Figure 2|Effects of H2S on dopaminergic neurons in Parkinson’s disease mouse model.

Figure 3|H2S promotes cell proliferation in the SVZ after MPTP-induced damage.
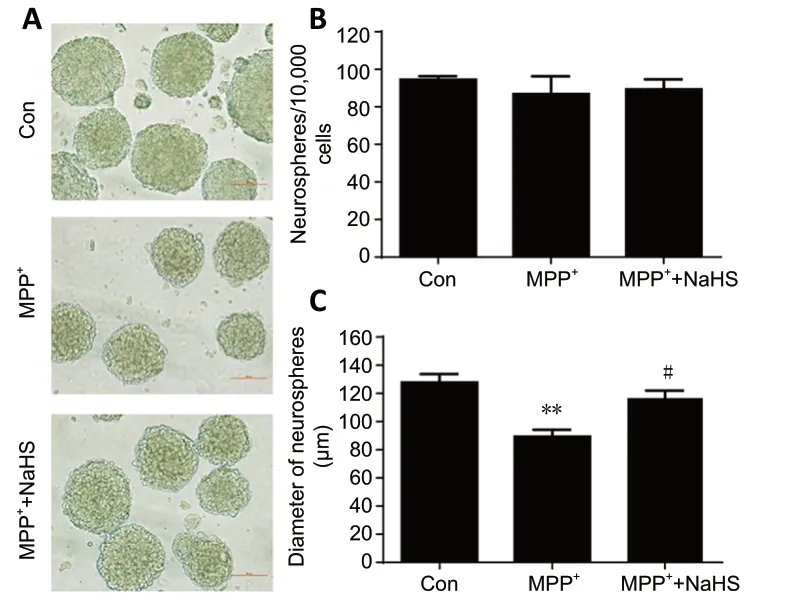
Figure 4|H2S relieves suppression of MPP+-induced adult neurogenesis in vitro.

Figure 5|H2S restores BrdU expression in MPP+-induced primary neurospheres.
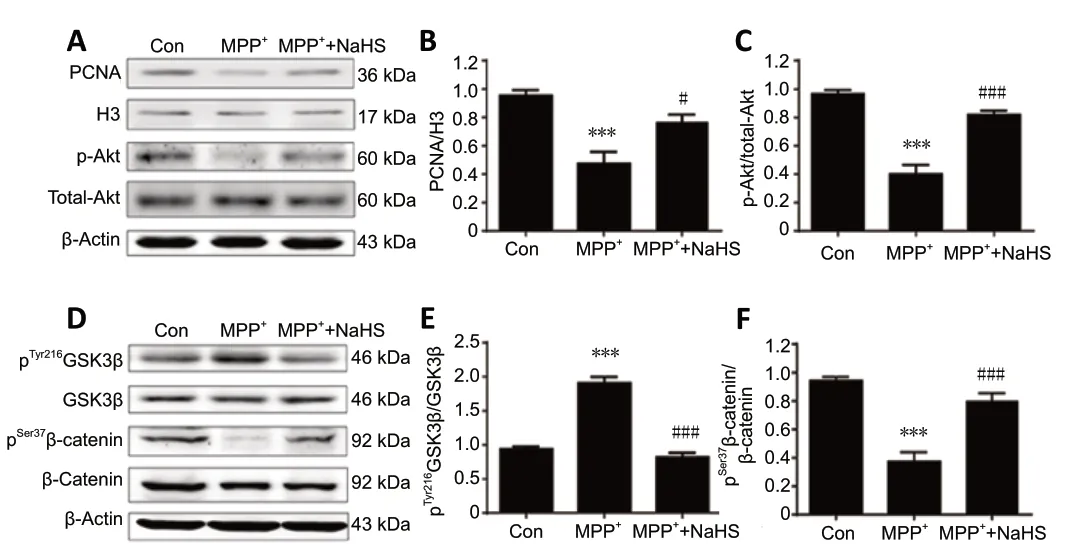
Figure 6|H2S enhances neurogenesis by upregulating the Akt/GSK-3β/β-catenin cascade in adult neural stem cells of the SVZ.
Financial support:The study was supported by the National Natural Science Foundation of China, Nos. 81803505 (to CQ), 81803498 (toLXW); the Natural Science Foundation of Jiangsu Province of China, No.BK20170564 (to CQ), and Yong Talents Training Program of Jiangsu University of China, No. 5521470000 (to JY). The funders had no roles in the study design, conduction of experiment, data collection and analysis,decision to publish, or preparation of the manuscript.
Institutional review board statement:The study was approved bythe Animal Care and Use Committee of Nanjing Medical University,China (IACUC Approval No. 1601153-3) on August 6, 2016. The animal experiment was reported according to the requirements of Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix,tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Fabricio Ferreira de Oliveira, Elysian Clinic, Brazil;Endre Pal, University of Pécs, Hungary.
Additional files:
Additional Figure 1:H2S promotes the expression of Ki67 in SVZ after MPTP-induced injury, but no significant effects on the expression of Ki67 in NaHS treatment alone.
Additional file 1:Open peer review reports 1 and 2.
Additional file 2:Original data of the experiment.

- 中国神经再生研究(英文版)的其它文章
- Clusterin: a multifaceted protein in the brain
- Positive effects of music therapist’s selected auditory stimulation on the autonomic nervous system of patients with disorder of consciousness: a randomized controlled trial
- Transcranial pulse current stimulation improves the locomotor function in a rat model of stroke
- Comparative transcriptomic analysis of rat versus mouse cerebral cortex after traumatic brain injury
- Delayed atomoxetine or fluoxetine treatment coupled with limited voluntary running promotes motor recovery in mice after ischemic stroke
- Extremely low frequency electromagnetic fields promote cognitive function and hippocampal neurogenesis of rats with cerebral ischemia