
外源赤霉素信号对甘蔗分蘖及其内源激素的影响
2021-12-08 06:26黎正英,丘立杭,闫海锋,周慧文,陈荣发,范业赓,梁和,吴建明
热带作物学报 2021年10期
黎正英,丘立杭,闫海锋,周慧文,陈荣发,范业赓,梁和,吴建明
摘 要:前期研究發现,外源GA信号影响了甘蔗分蘖进程。为了解GA信号对甘蔗不同分蘖时期叶片内源激素含量的影响,以综合性状优良的甘蔗主栽品种‘桂糖42号为材料,采用不同外源GA信号调节剂对蔗种进行浸种处理,并以清水处理为对照(CK),分别测定分蘖初期、盛期和末期+1叶(主苗+分蘖苗)中内源激素吲哚乙酸(indole-3-acetic acid,IAA)、赤霉素(gibberellins,GA)、乙烯(ethylene,ETH)、细胞分裂素(cytokinin,CTK)、脱落酸(abscisic acid,ABA)和油菜素内脂(brassinosteroids, BR)的含量,并分析它们之间的相关关系。结果表明:外源GA3(gibberellic acid 3;GA生物合成促进剂)对蔗种浸种处理后,增加了分蘖期甘蔗分蘖叶片中IAA、GA和CTK的总体含量,提高了分蘖前期ETH含量和后期ABA含量;多效唑(paclobutracol, PP333;GA生物合成抑制剂)处理后却增加了分蘖前期内源GA和ABA含量,提高了后期分蘖叶片中ETH含量,同时降低了CTK和BR含量。进一步的相关性分析发现,利用外源赤霉素信号促进剂处理蔗种后,甘蔗叶片内源激素IAA与GA、CTK和BR、CTK与GA均呈显著正相关;而ABA与GA呈显著负相关,与ETH呈显著正相关。可见,外源GA信号影响了甘蔗分蘖期叶片内源激素系统,促进生长的激素与抑制生长的激素达到动态平衡状态,最终导致了甘蔗分蘖的差异。甘蔗分蘖与植物内源激素含量密切相关,可通过施用外源激素来影响内源激素的含量,调控激素之间的动态平衡,达到提高甘蔗分蘖形成及其成茎的目的。
关键词:甘蔗;植物生长调节剂;赤霉素;多效唑;浸种;分蘖;内源激素
中图分类号:S533 文献标识码:A
Effect of Exogenous Gibberellin Signal on Sugarcane Tillering and Its Endogenous Hormones
LI Zhengying1, QIU Lihang2, YAN Haifeng2, ZHOU Huiwen2, CHEN Rongfa2, FAN Yegeng2, LIANG He1*, WU Jianming2*
1. College of Agriculture, Guangxi University, Nanning, Guangxi 530005, China; 2. Sugarcane Research Institute, Guangxi Academy of Agricultural Sciences / Sugarcane Research Center, Chinese Academy of Agricultural Sciences / Key Laboratory of Sugarcane Biotechnology and Genetic Improvement Guangxi, Ministry of Agriculture and Rural Affairs / Guangxi Key Laboratory of Sugarcane Genetic Improvement, Nanning, Guangxi 530007, China
Abstract: Previous studies found that the exogenous GA signal affected sugarcane tillering process. In order to understand the effect of GA signal on the content of endogenous hormones in leaves at different time of tillering stage of sugarcane, this experiment used the main sugarcane variety ‘Guitang 42 with excellent comprehensive traits as the plant material, and different exogenous GA signal regulators were used to soak the seedcanes with water treatment as the control (CK). The endogenous hormones indole-3-acetic acid (IAA) and gibberellins (GA), ethylene (ETH), cytokinin (CTK), abscisic acid (ABA) and brassinosteroids (BR) content in +1 leaves (main seedling + tiller seedling) at the early, full and late tillering stages were analyzed and the correlations between them were statistically tested. The results showed that after soaking sugarcane seedcanes with exogenous GA3 (Gibberellic acid 3; GA biosynthesis promoter), the total contents of IAA, GA and CTK in the tiller leaves was increased at the tillering stage, and the ETH content at the early stage and the ABA content at late tillering stage were increased. Paclobutracol (Paclobutracol, PP333; GA biosynthesis inhibitor) treatment increased the endogenous GA and ABA contents before tillering, and the ETH content at the later tiller leaves, but reduced the CTK and BR contents. Further correlation analysis found that in the treatment with exogenous gibberellin signal enhancer, the endogenous hormones of sugarcane leaves IAA and GA, CTK and BR, CTK and GA showed a significant positive correlation while ABA showed a significant negative correlation with GA but a significant positive correlation with ETH. The exogenous GA signal affected the endogenous hormone system in the leaves of sugarcane at the tillering stage, and the growth-promoting hormones and the growth-inhibiting hormones reached a dynamic equilibrium state, which ultimately led to the difference in the tillering. It is concluded that sugarcane tillering is closely related to the contents of plant endogenous hormones. Exogenous hormones could be used to influence the contents of endogenous hormones, regulate the dynamic balance between hormones, and achieve the purpose of improving sugarcane tiller formation and its stem formation.
Keywords: Saccharum officinarum; plant growth regulator; gibberellin; paclobutrazol; seedcane soaking; tiller; endogenous hormones
DOI: 10.3969/j.issn.1000-2561.2021.10.026
甘蔗(Saccharum officinarum)是一种蔗糖的重要原材料,并广泛产于世界热带、亚热带地区,属高光合效率C4作物。分蘖是甘蔗生长的一个重要特性,它直接影响甘蔗有效茎数,从而影响单位面积的产量。甘蔗产量=总有效茎数×平均单茎重,其中最终的总有效茎数取决于甘蔗的主茎和分蘖成茎,因此促进甘蔗分蘖,进而提高分蘖发育成有效茎的比例数,是提高甘蔗单产的关键[1-4]。
甘蔗分蘖的形成可分为分蘖芽的形成和分蘖芽的伸长生长,分蘖芽的伸长生长过程受到多种因素的影响和调控,它不仅受各种外界环境影响,如温度、光照、湿度以及水肥等,也受多种内源激素的共同调控,包括生长素(indole-3-acetic acid,IAA)、细胞分裂素(cytokinin,CTK)、赤霉素(gibberellins,GA)、脱落酸(abscisic acid,ABA)、乙烯(ethylene,ETH)、油菜素内酯(brassinosteroids,BR)等,内源激素之间的含量及平衡对分蘖发育进程至关重要[5]。
植物生长调节剂能够通过信号转导影响内源激素的含量或出现的时间来调节和控制植物的生长发育及各种生理过程,例如可直接或间接调控植物萌发、开花、生长等过程[6-7]。植物生长调节剂的作用效果与药剂种类密切相关,目前,在甘蔗上常用的主要植物生长调节剂有赤霉素类(gibberellins,GAs)、多效唑(paclobutrazol,PP333)和乙烯利(ethephon,ETP)等,它们在甘蔗产量形成和糖分积累中发挥了重要作用[8-9]。迄今,赤霉素类植物生长调节剂在园艺和农业生产上被广泛应用,并在推动作物增产和提质增效中做出了积极贡献[10]。赤霉素在甘蔗上的应用研究也有不少报道,邵廷富[11]和廖芬[12]研究发现,外源GA信号处理可以促进甘蔗茎秆伸长,增加节间长度;Jain等[13]利用GA3处理刺激了蔗芽生长,不仅提高了萌芽率,还增加了甘蔗株高和茎重;而范业赓等[14]利用高浓度GA3处理蔗种24 h后种植,结果却抑制了甘蔗的出苗和分蘖,进而导致甘蔗产量显著低于对照。多效唑是一种通过外源信号转导阻碍GA生物合成的抑制剂,不仅能够抑制茎秆的伸长[15],还能够促进作物分蘖并影响其生长发育进程[16-17],提高作物产量[18]等,这可能是多效唑通过信号转导抑制GA合成从而加速芽的分化进程所致[19]。由此可见,GA信号在甘蔗分蘖形成及其发育进程中扮演着重要角色。
众所周知,植物生长调节剂是人工合成的对植物的生长发育具有调节作用的化学物质,可作为植物内源激素合成与代谢的重要外源信号物质。赤霉素的主要生理效应是促进植物的茎秆伸长,尤其体现在节间伸长上。虽然赤霉素对甘蔗重要农艺性状的影响已有不少的相关研究[20],但赤霉素通路对甘蔗分蘖形成和发育的影响尚未可知,且在外源赤霉素信号影响甘蔗分蘖过程中内源激素间的平衡与含量变化关系的相关研究也较少。本研究以不同植物生长调节剂作为外源赤霉素信号,结合甘蔗叶蘖共生的特点,分析不同赤霉素信号浸种处理后甘蔗分蘖期叶片中内源激素的变化及其相关关系,为促进分蘖形成和调节分蘖成茎比例的甘蔗化学调控栽培提供技术支撑,也为甘蔗分蘖调控机理研究提供参考依据。
1 材料与方法
1.1 材料
供试甘蔗品种为主栽优良品种‘桂糖42号(GT42),种植于广西南宁市隆安县丁当镇广西农业科学院甘蔗研究所甘蔗繁育基地。
1.2 方法
试验设计2个不同赤霉素信号调节剂处理:赤霉素(GA3)和多效唑(PP333),清水处理为对照(CK),各调节剂处理浓度均为200 mg/L。将砍成双芽段的GT42种茎全部浸沒于不同赤霉素信号调节剂药液中,浸泡24 h后下种。
试验采取随机区组排列,重复3次,小区为5行,行长7 m,行距1.2 m,小区面积42 m2。试验材料于2016年3月10日种植,下种量为6000芽/hm2,其他参照王伦旺等[21]的栽培方法进行管理,并调查记录分蘖期各项农艺指标。
在甘蔗分蘖初期、盛期、末期,分别取样测定不同处理下不同分蘖期+1叶(主苗+分蘖苗)内源激素生长素吲哚乙酸(IAA)、赤霉素(GA)、乙烯(ETH)、细胞分裂素(CTK)、脱落酸(ABA)和油菜素内脂(BR)等的含量。内源激素提取参照陈明辉等[22]提取方法,其测定采用酶联免疫吸附分析法(enzyme-linked immunosorbent assay,ELISA),在6个植株中分别收集主苗和分蘖苗的+1叶样品,并混合形成一个生物重复样品,每个样品重复测定3次,相应激素测定试剂盒由中国农业大学作物化学控制实验室提供。
1.3 数据处理
采用Excel 2010软件进行数据编辑,SPSS 20.0软件进行LSD方差分析,GraphPad Prism 8.0软件绘图,图中的数据均为均值±标准差。
2 结果与分析
2.1 不同赤霉素信号调节剂处理后甘蔗分蘖生长情况
田间调查发现,不同赤霉素信号调节剂处理后蔗种萌发出苗的时间相当,但随着生育进程推进至分蘖期,甘蔗分蘖情况明显不同。GA3处理下甘蔗分蘖进程受到延缓和抑制,即分蘖苗破土时间比清水对照的晚了约7 d,且分蘖苗的生长也受主苗的抑制;然而PP333处理下的结果恰好相反,不仅提早促进了甘蔗分蘖的形成,且分蘖苗与主苗的生长几乎相同,即主苗的生长并未抑制分蘖苗的生长。由此可见,外源GA信号影响了甘蔗分蘖形成及其在分蘖期的生育进程。
2.2 不同赤霉素信号调节剂处理对甘蔗分蘖期叶片内源激素含量的影响
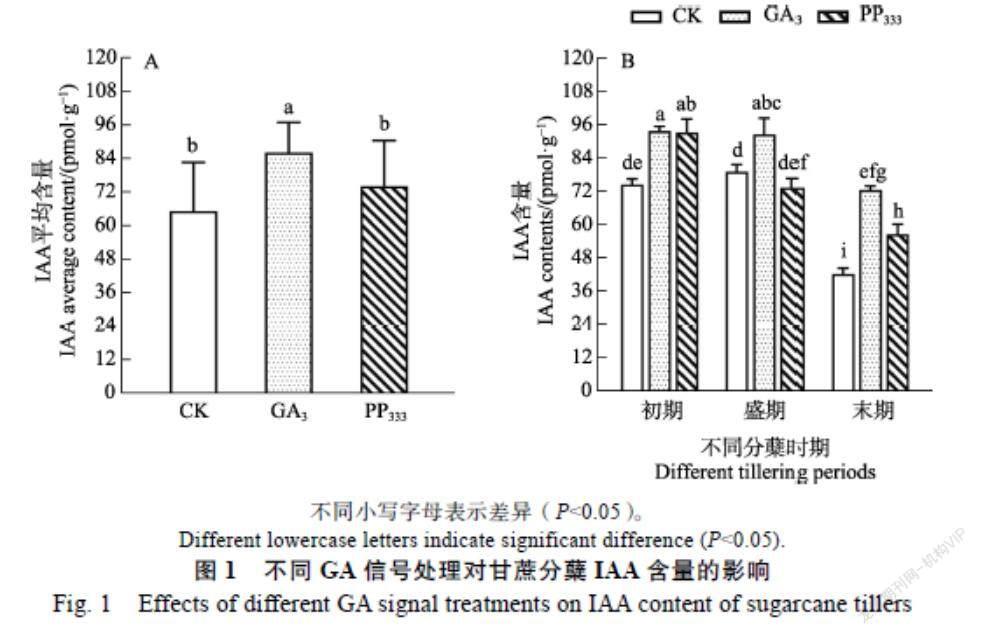
2.2.1 IAA 从图1可知,甘蔗不同分蘖期叶片的IAA在响应不同GA信号调节剂处理上存在差异。由图1A可知,在整个分蘖期,GA3处理和PP333处理总体上均增加了甘蔗分蘖期叶片中IAA的含量,且GA3处理后IAA的增加更加显著,这说明GA3和PP333处理都诱导了分蘖期IAA的积累,结合甘蔗分蘖情况,这暗示了GA3处理下IAA在主苗的积累可能是分蘖苗生长受抑制的原因。由图1-B可知,在甘蔗分蘖的不同时期,IAA含量在不同GA信号处理下也有明显的变化。在分蘖初期,GA3处理和PP333处理的IAA含量都显著高于对照。当到了分蘖盛期,GA3处理后IAA的含量仍显著高于对照的水平,但PP333处理下的含量反而低于对照,这暗示分蘖盛期叶片中IAA的持续积累不利于分蘖的形成和发育。当分蘖进入末期之后,新生分蘖骤减或者无分蘖,此时不同处理下叶片中的IAA含量比初期和盛期的明显下降,但GA3处理和PP333处理下的IAA含量仍显著高于对照水平,且此时GA3处理的IAA含量与对照的初期和盛期的水平相当,这些结果进一步说明GA3抑制和延缓甘蔗分蘖与IAA的含量息息相关。
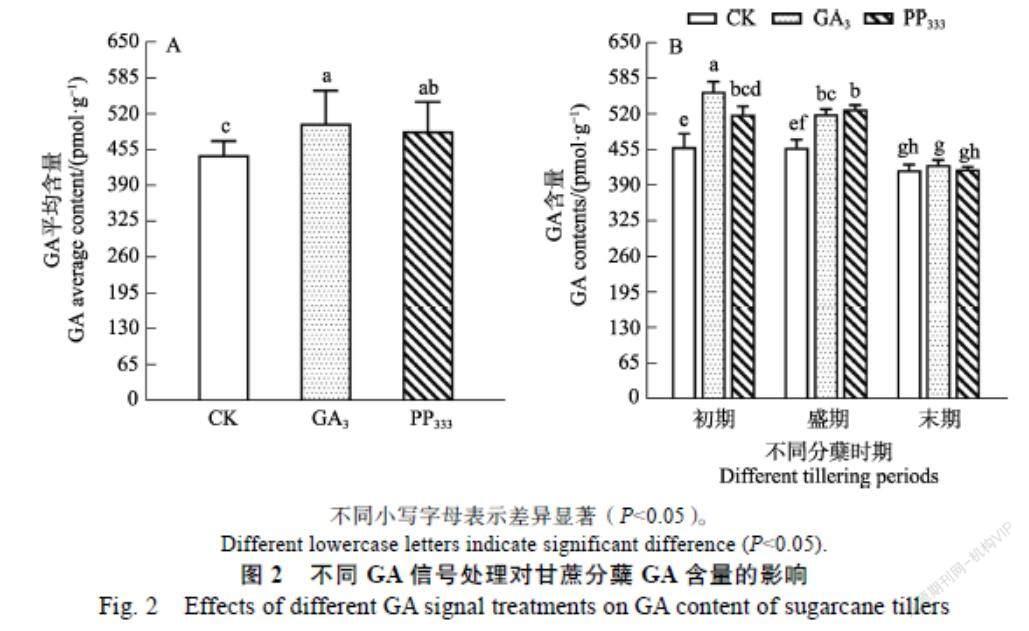
2.2.2 GA 从图2可知,不同GA信号调节剂浸种处理后,对甘蔗不同分蘖期叶片中的GA含量有不同影响。由图2A可知,在整个分蘖期,GA3处理和PP333处理的分蘖期叶片中GA含量都显著高于对照,且GA3处理的更加显著,这说明GA3处理和PP333处理都诱导了分蘖期GA的表达。在甘蔗的不同分蘖时期,不同GA信号处理下的GA含量也有所差异。由图2-B可知,在分蘖初期,GA3处理和PP333处理下分蘖期叶片中的GA含量都显著高于对照,这暗示分蘖初期GA含量的升高有利于分蘖芽的萌动。随着分蘖期的推进,在分蘖盛期时,GA3处理下的GA含量下降,而PP333处理仍处于高水平,这说明持续高水平的GA含量有利于分蘖芽的生长发育,这暗示在分蘖盛期GA3处理下主苗中GA含量的下降可能是外源GA3信号延缓甘蔗分蘖的原因。当进入分蘖末期,两个处理下的GA含量与对照无明显变化,说明在分蘖末期GA含量的降低有利于分蘖苗生长。
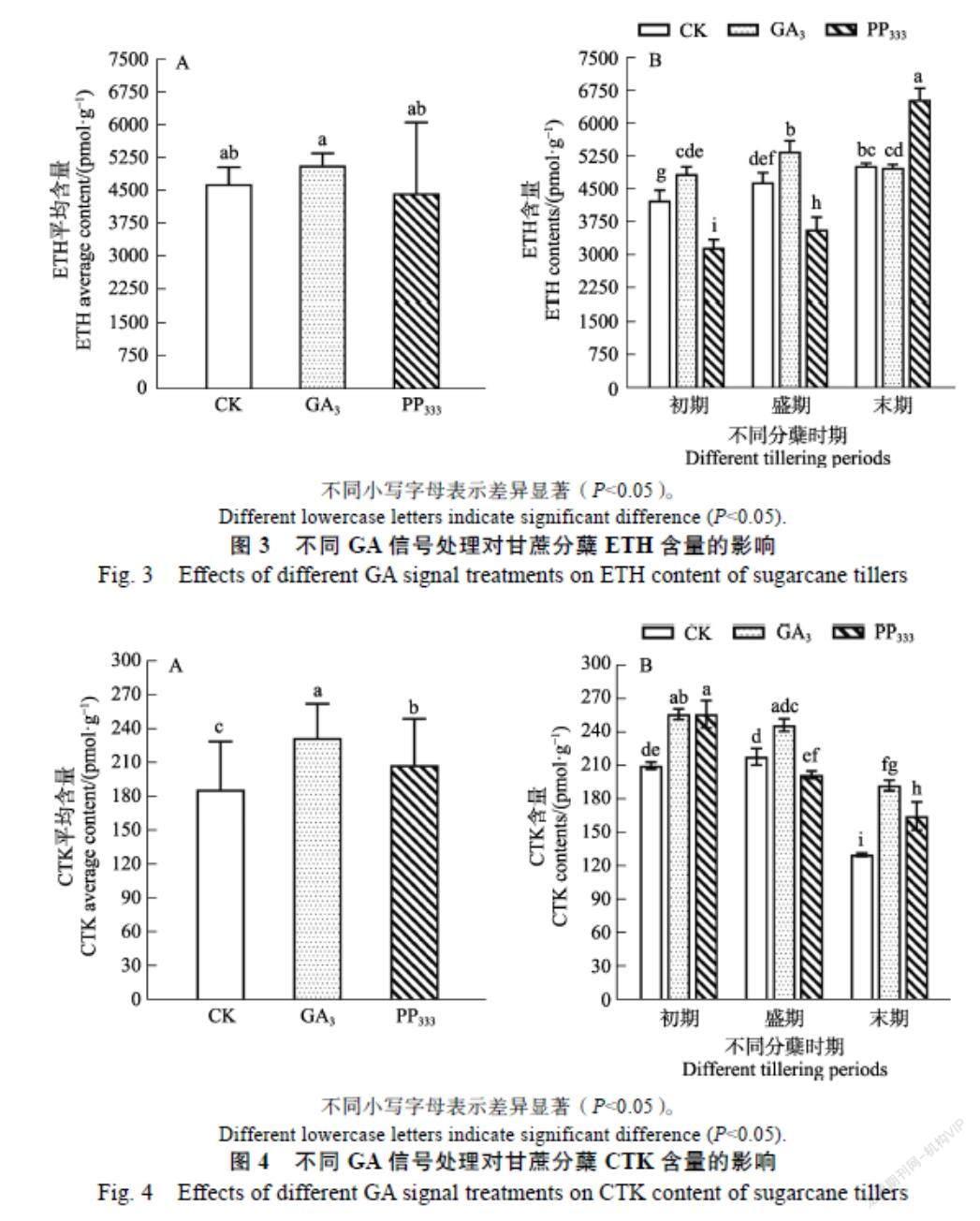
2.2.3 ETH 从图3可知,不同GA信号调节剂处理对甘蔗分蘖期叶片中的ETH含量存在不同影响。从图3A可知,在整个分蘖期,GA3处理的分蘖叶片中ETH含量高于对照,而PP333处理的结果相反。在甘蔗不同分蘖时期,不同GA信号调节剂处理下,分蘖期叶片中的ETH含量也发生了明显的变化。从图3B可知,在分蘖初期,GA3处理的ETH含量显著高于对照,而PP333处理下却显著低于对照,这暗示了ETH含量的升高制约着分蘖芽的萌动。当进入分蘖盛期后,PP333处理的ETH含量极显著低于对照,而GA3处理显著高于对照,说明分蘖盛期叶片中低浓度ETH的释放有利于甘蔗分蘖,这暗示高水平的ETH含量可能是外源GA3延缓甘蔗分蘖芽生长的重要因素之一。而到了分蘖末期,GA3处理的ETH含量下降到与对照水平相当,说明此时GA3处理下的分蘖开始增多,PP333处理下的分蘖已不生长,这些暗示了外源激素GA信号诱导ETH的增加不利于分蘖的生长发育及成苗。
2.2.4 CTK 从图4可知,不同GA信号调节剂处理对甘蔗分蘖期叶片的CTK含量的有所差异。从图4A可知,GA3处理和PP333处理对甘蔗分蘖期叶片的CTK含量的积累效果是显著的。从图4B可知,在甘蔗不同分蘖时期,分蘖期叶片的CTK含量在不同GA信号调节剂处理下发生了明显的变化。在分蘖初期,GA3处理和PP333处理的CTK含量显著高于对照,这说明在分蘖初期CTK含量的增加有利于分蘖芽的萌动。而到了分蘖盛期,PP333处理的CTK含量下降且显著低于对照,GA3处理则与初期无明显变化,这说明在分蘖盛期低水平的CTK含量对甘蔗分蘖芽的分化和生长是有益的。随着甘蔗分蘖的推进,到了分蘖末期,PP333處理的CTK含量升高,但仍显著低于GA3处理,这暗示了GA3处理下的保持高水平的CTK含量也可能是延缓甘蔗分蘖的重要因素之一。
2.2.5 ABA 从图5可知,不同GA信号调节剂处理对甘蔗分蘖期叶片的ABA含量影响不同。从图5A可知,GA3处理和PP333处理下分蘖期叶片的ABA含量总体高于对照,说明这两个处理对甘蔗分蘖期叶片的ABA积累是有益的。从图5B可知,在甘蔗不同分蘖时期,叶片的ABA含量在不同GA信号调节剂处理下也有明显的变化。在分蘖初期,GA3处理的ABA含量受到了抑制,显著低于对照,而PP333处理下的ABA含量则与之相反。说明PP333诱导了ABA的表达,GA3则相反,这暗示在分蘖前期时,ABA含量的下降对分蘖芽的萌动有很大影响。而到了分蘖盛期,GA3处理反而诱导了ABA的合成,ABA含量不断增幅,显著高于对照,PP333处理下的ABA含量反而下降且显著低于对照。说明随着分蘖的推进,ABA含量的降低有利于分蘖形成和生长。当分蘖进入末期,不同处理下的ABA含量整体升高,PP333处理下的ABA含量显著高于GA3处理和对照,这暗示了GA3抑制和延缓主苗与分蘖苗生长可能与分蘖盛期叶片中ABA含量的积累有关。
2.2.6 BR 从图6可知,不同GA信号调节剂浸种处理下,甘蔗分蘖期叶片的BR含量有所差异。从图6A可知,在整个分蘖期,GA3处理能有效地积累分蘖期叶片的BR含量。从图6B可知,不同GA信号处理下,不同分蘖时期的BR含量也有明显变化。在分蘖初期,GA3处理和PP333处理对甘蔗分蘖期叶片的BR含量影响不大。当进入分蘖盛期时,GA3处理和PP333处理下的BR含量显著高于对照,说明这两个处理都促进了BR的合成,这暗示了分蘖盛期叶片中BR含量的积累
有利于分蘖的形成和生长发育。而进入分蘖末期后,PP333处理的BR含量与对照无明显差异,GA3处理却极显著高于对照,这暗示GA3处理下BR在主苗中的持续积累可能是延缓和抑制分蘖苗生长的重要因素。
2.3 甘蔗分蘖率与内源激素之间的相关关系
如表1所示,甘蔗分蘖率与内源激素含量存在一定的相关性,分蘖期叶片中不同内源激素含量之间也存在着不同程度的差异。在甘蔗分蘖期,分蘖率与内源激素含量的相关系数表现均没有达到显著水平,而甘蔗叶片内源激素IAA与GA、CTK和BR、CTK与GA呈显著正相关;ABA与GA呈显著负相关,与ETH呈显著正相关。
在分蘖期,IAA与GA呈显著正相关,结合图1和图2分蘖期内源激素的含量分布情况,进一步说明了外源GA信号延缓或抑制甘蔗分蘖可能与IAA和GA的含量协同增高有关。IAA和CTK也有协同关系,结合图1和图4可知,在GA抑制分蘖的情况下,IAA与CTK的比值增大是确保甘蔗分蘖的基础。BR具有促进作物生长的生理功能,BR与IAA呈显著正相关,结合图6可知,虽然外源GA3延缓了甘蔗的分蘖,但有BR存在的情况下,并没有抑制分蘖生长,这说明分蘖叶片中BR的存在是GA3抑制分蘖情况后仍能保障分蘖的内在因素之一。
GA与CTK也呈正相关,这进一步说明IAA与GA、IAA与CTK之间的协同关系,它们之间相互作用共同调控甘蔗节间的生长发育。而GA和ABA呈显著负相关,二者通过拮抗作用调控甘蔗体内GA与ABA含量,结合图2和图5可知,GA与ABA的比值动态影响着分蘖芽的萌动及形成。ABA和ETH呈显著正相关,结合图3和图结果5可知,随着甘蔗分蘖的增多,外源GA信号诱导了ABA信号增强,同时叶片ETH的释放量也随之增多,而外源GA信号又延缓了分蘖的发生和生长,这暗示ABA和ETH含量协同增高是外源GA信号延缓或抑制分蘖进程的重要原因之一。
3 讨论
3.1 GA信号延缓甘蔗分蘖的形成与进程
植物生长调节物质是通过改变植物体内内源激素系统来调节植物生长发育进程[8]。前人研究发现,GA信号代谢通路与甘蔗分蘖密切相关,独角金内酯(strigolactone,SL)的合成受GA信号的调控,SL的特异性受体D14蛋白能够和DELLA蛋白结合形成复合物,间接调控水稻分蘖[23],而DELLA蛋白是GA信号转导途径的核心元件,属于GRAS蛋白[24],由此可见,GA信号系统对分蘖的进程发挥了重要作用。
本研究结果也表明,利用外源赤霉素信号促进剂处理蔗种后,甘蔗分蘖期叶片内源激素系统受到干扰,抑制和延缓了甘蔗分蘖的进程,而用赤霉素信号抑制剂处理,则促进了分蘖的提早发生且分蘖数目增多。由此可见,植物激素信号传导相关的代谢通路,在甘蔗分蘖的发生及生长发育进程中发挥了重要作用,尤其是GA信号传导。
3.2 内源激素含量变化及平衡影响甘蔗分蘖的形成与进程
植物内源激素在植物生长发育的各个过程中发挥着重要的调控作用,植物可通过控制体内激素的含量及其平衡来影响甘蔗分蘖的发生和成茎[25]。前人研究发现,IAA通过顶端优势促进主茎生长,同时抑制侧芽的生长发育[26]。而CTK与IAA产生的作用相反,CTK作为一种促进信号能够解除侧芽生長的抑制。研究表明,外源增施氮肥可以提高分蘖芽的CTK含量,从而显著诱导植株分蘖芽的萌发[27]。本研究结果也表明,在外源GA信号处理下,分蘖叶片中IAA含量的持续积累不利于分蘖的形成和发育,而在分蘖初期CTK含量的增加反而是有利于分蘖芽的萌动,这可能与CTK在分蘖前期解除了分蘖芽生长的抑制有关。
内源激素GA在打破种子休眠,促进萌动[28],促进节间伸长[29],抑制分蘖芽的发育[30]上发挥重要作用。本研究结果发现,在外源GA信号处理下,分蘖前期叶片中GA含量的增加促进了分蘖芽的萌发,而随着分蘖期的推进,GA含量的下降则不利于分蘖芽的生长发育,与前人研究结果一致。BR和GA有着相同的功能,如促进茎伸长,种子萌发,抑制侧芽生长发育[31-32]等。本研究中,在GA3处理下BR在主苗中的持续积累可能是延缓和抑制分蘖苗生长的原因。ABA能够抑制细胞生长、调控分蘖芽休眠、萌发和幼苗早期发育[33-34]。本研究结果表明,外源GA信号调控了甘蔗分蘖叶片中GA和ABA含量,GA和ABA含量比值调控分蘖芽的萌动和形成。
王威豪等[35]研究发现,在干旱条件下,在叶面喷施乙烯利能促进甘蔗分蘖芽的萌动,乙烯利浓度越高其效应越显著。而本研究结果发现,外源GA信号抑制乙烯的释放反而有利于甘蔗分蘖。本研究结果与前人的报道结果似乎相矛盾,这可能是由于品种差异、砍种时间、样品处理方式等不一致导致的。
3.3 内源激素之间互作及相互关系影响甘蔗分蘖的形成与进程
内源激素在甘蔗分蘖过程中存在复杂的相互关系,激素之间的平衡对甘蔗分蘖的生长生育有重要的调节作用。外源GA信号通过改变甘蔗分蘖叶片中内源激素IAA、GA、ETH、CTK、ABA和BR等含量及激素间的平衡来调节自身的代谢水平,从而影响甘蔗分蘖的形成与进程。
前人研究表明,外源GA信号能促进甘蔗叶片的内源激素IAA的合成,GA与IAA含量呈正相关[36],GA对CTK也有促进作用[37]。IAA和CTK在甘蔗分蘖芽的萌动和生长过程中具有重要作用[38-39]。研究发现,CTK可能是通过提高部分GA含量来促进分蘖生长[40]。本研究结果也表明,外源GA信号促进了IAA、GA和CTK的合成,IAA、GA与CTK之间存在协同关系,外源GA信号能延缓甘蔗分蘖可能与IAA和GA含量的协同增高有关,当甘蔗分蘖受GA抑制时,IAA与CTK的比值增大是确保甘蔗分蘖的基础。BR不仅在促进分蘖伸长方面具有效果显著,还能抑制生长素氧化酶的活性,提高植物内源IAA的含量,因此当BR与IAA含量同时增加时,能有效地提高甘蔗分蘖。本研究得出相似的结果,BR和IAA显著正相关,二者协同互作共同调控分蘖的生长发育。有研究表明,高水平BR对GA信号传导有抑制作用[41],BR和GA协同调控种子萌发进程[42]。本研究结果也表明,在BR存在的情况下,外源GA信号虽然抑制了甘蔗的分蘖,但分蘖的生长并没有受到延缓,说明在分蘖受到GA3抑制时分蘖叶片中BR的存在是仍能保障分蘖的内在因素之一。
植物内源激素GA和ABA具有拮抗作用,二者是调控分蘖成茎的关键因子,GA和ABA的比值高低控制着分蘖的发生[5]。本试验研究结果与前人报道一致,GA和ABA呈显著负相关,GA与ABA的比值动态影响着分蘖芽的萌动及形成。研究发现,ABA和独角金内酯(SL)也存在相互调控关系,SL与ABA合成信号途径紧密耦联共同调控水稻分蘖发育的分子机制[43],而独角金内酯的合成也受GA信号的调控[22],由此可见,在分蘖的生长发育进程中GA信号途径与ABA信号途径发挥了重要作用。本研究结果也发现,外源GA信号能诱导ABA信号增强,说明在分蘖的发育进程中ABA和GA信号之间协同调控分蘖成茎。外源GA信号在乙烯促进分蘖成茎中也起重要作用[44],研究发现外源GA促进甘蔗节间伸长可能是通过抑制ABA和ETH含量,同时提高GA3含量来调节节间伸长[20],本研究结果表明,在外源GA信号处理下,甘蔗分蘖叶片的ABA和ETH呈显著正相关,ABA和ETH含量的升高制约着分蘖芽的萌动,乙烯释放量的增加和ABA含量的增高可能是外源GA信号延缓或抑制分蘖进程的重要原因之一。
甘蔗分蘖与内源激素密切相关,但不同植物激素的影响程度存在差异,内源激素之间的关系也错综复杂,施以外源GA信号能影响分蘖期内源激素系统,最终导致分蘖的差异。这说明外源GA信号对甘蔗内源激素的影响是相互关联的,促进生长的激素与抑制生长的激素的平衡状态决定了甘蔗分蘖效果。
综上所述,甘蔗分蘖受多种植物内源激素的调控,激素间的互作共同调控分蘖的生长发育,可通过外源GA信号来影响甘蔗体内内源激素之间的平衡,达到提高甘蔗分蘖的形成和成茎的目的。
参考文献
[1] Esayas T, Firew M, Amsalu A. Correlation and path coefficient analyses in sugarcane genotypes of Ethiopia[J]. American Journal of Plant Sciences, 2016, 7(10): 1490-1497.
[2] 丘立杭, 范业庚, 罗含敏, 等. 甘蔗分蘖发生及成茎的调控研究进展[J]. 植物生理学报, 2018, 54(2): 192-202.
[3] 周慧文, 范业赓, 陈荣发, 等. 不同代数健康种苗分蘖差异及浸种处理对一代种茎繁育的影响研究[J]. 中国糖料, 2019, 41(1): 33-36.
[4] 罗含敏, 周慧文, 闫海锋, 等. 甘蔗产量形成的要素及其化控措施[J]. 广西糖业, 2020(4): 8-14.
[5] 丘立杭, 李 强, 黄 杏, 等. 弱光胁迫影响甘蔗叶片内源激素的平衡和分蘖进程[J]. 植物生理学报, 2017, 53(2): 280-290.
[6] 王 熹. 试论我国作物化控研究的发展[J]. 作物杂志, 1993(2): 1-4.
[7] 张 悦. 植物生长调节剂对作物调控效应的研究现状[J]. 现代化农业, 2016(5): 31-34.
[8] 李杨瑞. 现代甘蔗学[M]. 北京: 中国农业出版社, 2010: 1-4, 282.
[9] 陈迪文, 吴庚福, 周文灵, 等. 甘蔗糖分调控因素与增糖应用研究现状[J]. 甘蔗糖业, 2020(3): 43-51.
[10] 黎 家, 李传友. 新中国成立70年来植物激素研究进展[J]. 中国科学: 生命科学, 2019, 49(10): 1227-1281.
[11] 邵廷富. 赤霉素对甘蔗生长的影响[J]. 植物生理学通讯, 1965(4): 8-10.
[12] 廖 芬. 赤霉素、多效唑对甘蔗茎尖分生组织的解剖效应及与蔗茎增粗的关系[D]. 南宁: 广西大学, 2004.
[13] Jain R, Singh S N, Solomon S, 等. 赤霉素对甘蔗发芽和早期蔗茎生长的潜在调节作用(英文)[J]. 广西农业科学, 2010, 41(9): 1025-1028.
[14] 范业赓, 陈荣发, 周慧文, 等. 不同植物生长调节剂浸种对甘蔗分蘖及产量性状的影响[J]. 中国糖料, 2019, 41(2): 23-27.
[15] 许林英, 周南镚, 戎国增, 等. 植物生长调节剂对棉花基质育苗幼苗生长的影响[J]. 浙江农业科学, 2009(2): 325-327.
[16] 刘晓静, 张德罡. 多效唑和稀效唑对草地早熟禾分蘖和根的生长特性的影响[J]. 草原与草坪, 2005(6): 37-40.
[17] 刘俊仙, 李 松, 谭 芳, 等. 多效唑浸种对甘蔗分蘖提早萌发及幼苗内源激素含量的影响[J]. 湖南农业科学, 2016(2): 22-25, 30.
[18] 周建霞, 丰兆平, 陈晓阳, 等. 分蘖期施用多效唑对杂交水稻C两优817产量及主要性状的影响[J]. 杂交水稻, 2021, 36(2): 72-74.
[19] 金曉蕾, 刘景辉, 罗中旺, 等. 叶面喷施PP333和6-BA对甜荞花芽分化及内源激素的影响[J]. 西北植物学报, 2019, 39(11): 1988-1997.
[20] 吴建明, 李杨瑞, 杨 柳, 等. 赤霉素诱导甘蔗节间伸长与内源激素变化的关系[J]. 热带作物学报, 2009, 30(10): 1452-1457.
[21] 王伦旺, 廖江雄, 谭 芳, 等. 高产高糖抗倒伏甘蔗新品种‘桂糖42号的选育及高产栽培技术[J]. 南方农业学报, 2015, 46(8): 1361-1366.
[22] 陈明辉, 黄 杏, 谢晓娜, 等. 宿根矮化病菌对甘蔗生长和内源激素的影响[J]. 华中农业大学学报, 2013, 32(3): 57-61.
[23] Shinsaku I, Daichi Y, Mikihisa M, et al. Regulation of strigolactone biosynthesis by gibberellin signaling[J]. Plant physiology, 2017, 174(2): 1250-1259.
[24] 白云赫, 朱旭東, 樊秀彩, 等. 植物DELLA蛋白及其应答赤霉素信号调控植物生长发育的研究进展[J]. 分子植物育种, 2019, 17(8): 2509-2516.
[25] Li X, Qian Q, Fu Z, et al. Control of tillering in rice[J]. Nature, 2003, 422(6932): 618-21.
[26] Reinhardt D, Mandel T, Kuhlemeier C. Auxin regulates the initiation and radial position of plant lateral organs[J]. The Plant Cell, 2000, 12(4): 507-518.
[27] 许俊旭. 水稻分蘖芽萌发与休眠相互转换的激素调控和分子机制[D]. 南京: 南京农业大学, 2015.
[28] 粟莉圆, 苏丽华, 覃建彬, 等. 不同家系火力楠种子性状变异及外源赤霉素对种子萌发的影响[J]. 种子, 2020, 39(7): 49-52, 59.
[29] 吴建明, 李杨瑞, 王爱勤, 等. 赤霉素处理对甘蔗节间伸长及产质量的影响[J]. 中国糖料, 2010(4): 24-26.
[30] 洪晓富, 蒋彭炎, 郑寨生, 等. 水稻分蘖期喷施赤霉素(GA3)对控制分蘖和提高成穗率的效果[J]. 浙江农业科学, 1998(1): 3-5.
[31] 俞明宏, 王力明, 刘 继, 等. 表油菜素内酯对镉胁迫下番茄幼苗生长及镉累积的影响[J]. 中国土壤与肥料, 2020(3): 151-156.
[32] 侯会云. 油菜素内酯对盐胁迫下水稻种子萌发的影响[J]. 热带农业科学, 2020, 40(7): 1-6.
[33] 王光明, 刘保国, 陈 静, 等. 内源激素ABA对水稻再生芽萌发的影响[J]. 西南农业大学学报, 1997(4): 24-28.
[34] Miao C, Xiao L, Hua K, et al. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity[J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(23): 6058-6063.
[35] 王威豪, 王一丁, 莫云川, 等. 水分胁迫下喷施乙烯利对甘蔗分蘖及农艺性状的影响[J]. 广西农业科学, 2007(2): 148-151.
[36] 黄诚梅, 班德宇, 魏源文, 等. 赤霉素与多效唑对甘蔗愈伤再生苗植株内源激素含量的影响[J]. 热带农业科学, 2019, 39(4): 41-47.
[37] 张 晨, 周 涛, 郑 伟, 等. 外源赤霉素对太子参块根中细胞分裂素含量及其代谢关键酶基因表达的影响[J]. 南方农业学报, 2018, 49(11): 2141-2147.
[38] 周传凤, 李杨瑞, 杨丽涛. 乙烯利对甘蔗伤流液氮化物和钙含量的影响及其与分蘖的关系[J]. 广西农业科学, 2007(3): 258-262.
[39] 周传凤, 李杨瑞, 杨丽涛. 甘蔗分蘖期间叶面喷施乙烯利后两种内源激素的变化[J]. 西南农业学报, 2007(3): 388-391.
[40] 郑庆伟. 版纳植物园揭示赤霉素和细胞分裂素相互作用促进小桐子分枝生长分子机理[J]. 农药市场信息, 2017(26): 50-51.
[41] 张 慧. 油菜素内酯和HY5在低比例红光/远红光下诱导赤霉素调控番茄叶茎伸长的机制研究[D]. 杭州: 浙江大学, 2019.
[42] 熊 敏, 李钱峰, 刘巧泉. 油菜素内酯和赤霉素通过影响谷蛋白丰度调控水稻种子萌发的研究[C]//2018中国作物学会学术年会论文摘要集. 北京: 中国作物学会, 2018: 1.
[43] Liu X, Hu Q, Yan J, et al. ζ-Carotene isomerase suppresses tillering in rice through the coordinated biosynthesis of strigolactone and abscisic acid[J]. Molecular Plant, 2020, 13: 1784-1801.
[44] 叶燕萍. 乙烯利促进甘蔗有效分蘖的生理生化机理研究[D]. 南宁: 广西大学, 2006.
责任编辑:白 净
猜你喜欢
农学学报(2022年4期)2022-07-14
农民致富之友(2019年9期)2019-05-22
农家科技(2018年2期)2018-05-05
新农业(2017年6期)2017-07-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
现代农业科技(2016年20期)2016-12-20
新农村(2015年10期)2015-09-28
安徽农学通报(2015年10期)2015-06-15
山西果树(2014年3期)2014-07-15
