
水杉原生种群胸径树高与树冠的通径分析1)
2021-12-03 07:13赵保国朱江艾训儒姚兰郭秋菊洪建峰
东北林业大学学报 2021年10期
赵保国 朱江 艾训儒 姚兰 郭秋菊 洪建峰
(湖北民族大学,恩施,445000) (湖北星斗山国家级自然保护区管理局)
水杉(Metasequoiaglyptostroboides)是我国特有的世界著名的杉科水杉属孑遗植物,被誉为植物界的“活化石”,一度被认为已经全部灭绝,直到20世纪40年代在我国湖北利川谋道镇被发现[1]。水杉的发现被公认为是我国甚至世界20世纪植物界的重大发现[2-3]。水杉从发现至今已有70多年,一直受到国内外学者们极大的重视,研究表明水杉原生种群呈衰退型[4-6],原生环境林下鲜有幼苗发现,存在天然更新困难[7-9],影响水杉种群天然更新的因素存在种源限制和种子扩散限制[9]。种子扩散过程除了受扩散方式的制约,还受母体特征的特性制约[10],而树冠作为水杉球果的生长发育场所,冠幅大小和冠形等直接影响着水杉原生母树的种子产量和林下植被的生活型,影响了种子的着地几率、着地后的环境,与水杉的天然更新等存在密切联系。树冠作为林木的重要组成部分[11],是其与周围生长环境相互作用的结果,是林木生理过程的主要场所,对林木的生长过程具有主导地位[12-13]。然而,现有的关于水杉原生种群的研究较少涉及树冠层。
通径分析是数量遗传学家Sewall Wright[14]在1921年提出来的一种多元统计技术,是简单相关分析的继续,在多元回归的基础上将相关系数加以分解,通过直接通径、间接通径表示某一变量对因变量的直接作用效果、通过其他变量对因变量的间接作用效果[15]。胸径、树高、冠幅等是树木的重要生理指标,有许多学者对柚木[16]、樟子松[17-18]、华北落叶松[19]、杉木[12,15,20]、秃杉[11]等对象开展了类似的研究。本文选取胸径、树高、冠幅等生理指标,利用通径分析[15,21]研究水杉原生种群,明确水杉原生种群各影响因子与树冠之间的关系,充分考虑各指标对水杉原生种群树冠的直接影响和间接影响。研究表明,不同林龄的林分树冠存在较大的差异[22]。目前,国内外尚未对水杉原生种群的树冠特性进行详细分析,本文以星斗山国家级自然保护区内的水杉原生种群为研究对象,分析和阐明水杉原生种群不同林龄个体的胸径、树高和树冠之间的联系,为进一步建立胸径、树高、树冠的精准模型提供依据,为水杉原生种群的形态特征和个体生长发育的研究提供理论支撑,为水杉原生种群的保护和天然更新提供参考。
1 研究区概况
星斗山国家级自然保护区分为东、西两个片区,区域内年平均气温12 ℃,年平均降水1 481 mm,四季分明,雨热同期,为典型的亚热带大陆性季风气候。水杉原生种群主要分布于保护区的西区(108°31′~108°48′E、30°4′~30°14′N),海拔范围830~1 565 m,区域内主要为黄棕壤和黄壤,水资源充足,独特的地理条件形成了水杉原生母树的避难所。
2 研究方法
2.1 水杉原生种群个体性状调查
2017—2018年间对保护区内全部水杉原生种群(5 663株)进行调查,使用胸径尺测量胸径,使用激光测距仪测量树高和活枝下高,利用皮尺和激光测距仪测量冠幅长度(南北、东西)。
2.2 径级与林龄划分
通过径级代替龄级[5,23]的方法,结合水杉径级分布将水杉原生种群分为3个年龄阶段,分别为中龄树(小于100 a),成年树(100~200 a)和老龄树(大于200 a)。
2.3 树冠因子的计算
冠幅(W):用东西向冠幅和南北向冠幅的平均值表示树木的冠幅长度,其计算公式为:
W=(W1+W2)/2。
式中:W1、W2表示东西向与南北向的冠幅长度(m)。
冠长(L):指树木顶部到第一个活枝的长度。其计算公式为:
L=H-H1。
式中:H为树高(m);H1为树干第一个活枝高度(m)。
冠长率(R):指树冠长度在树干中所占的比率。具体公式为:
R=L/H。
冠形率(S):冠形率用冠长(L)与冠幅(W)的比值表示:
S=L/W。
以3 m为级距对冠幅进行分级,18 m为分级上界,共分为7级,从而使不同林龄的水杉原生母树冠幅占比更好体现,同理,采用类似冠幅分级的方式将冠长率以0.1为级距分为10级。
参照杜家菊[24]、宋小园[25]等,使用SPSS对数据进行处理分析,根据通径分析理论和计算公式,得出直接通径系数、间接通径系数和决策系数。决策系数是通径分析中的决策指标,用它可以把自变量对响应变量的综合作用进行排序,确定主要决策变量和限制变量[26]。其计算公式具体为:
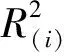
3 结果与分析
3.1 胸径和树高结构
水杉原生种群不同林龄阶段的胸径、树高的统计信息见下表。由表1、表2可知,星斗山自然保护区水杉原生种群中,胸径变化在24.2~248.0 cm,中龄树、成年树、老龄树的平均胸径依次为60.8、72.9、98.1 cm,随着林分年龄增大,胸径增大的趋势也在发生变化,其中,平均胸径在不同年龄阶段的增长率依次为19.90%、34.60%。通过表3可知,水杉原生母树的树高主要分布在9~51 m,中龄林、成年林、老龄林的平均树高为25.4、29.3、36.0 m,树高在不同年龄阶段的增长率为15.35%、22.80%,但最大值出现在成年树。通过差异性分析表明,不同林龄水杉原生母树的平均胸径与平均树高均具有显著性差异(p<0.01)。

表1 水杉原生种群胸径分布
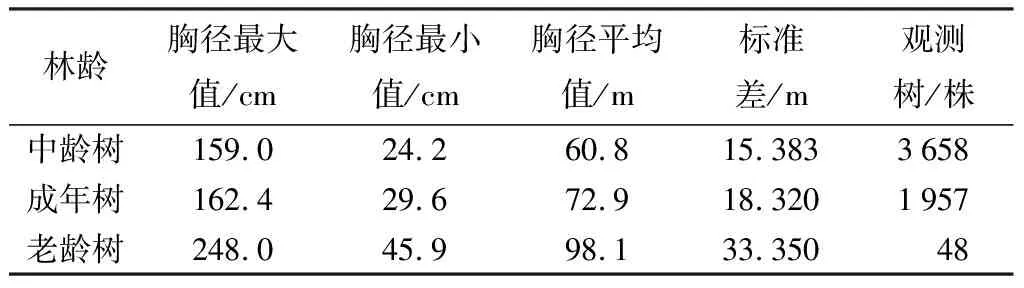
表2 水杉原生种群不同林龄胸径统计

表3 水杉原生种群不同林龄树高统计
3.2 树冠结构
通过表4、表5可知,平均冠长和冠幅都在随着水杉年龄的增加而增大,中龄林、成年林、老龄林的平均冠长依次为21.2、24.6、30.2 m,水杉冠长在各个阶段的增长率分别为16.04%、22.93%。水杉原生母树的平均冠幅在各个阶段依次为7.7、8.9、10.6 m,冠幅的变化率依次为15.58%、19.09%,不同林龄的水杉冠幅和冠长都具有显著差异(p<0.01)。
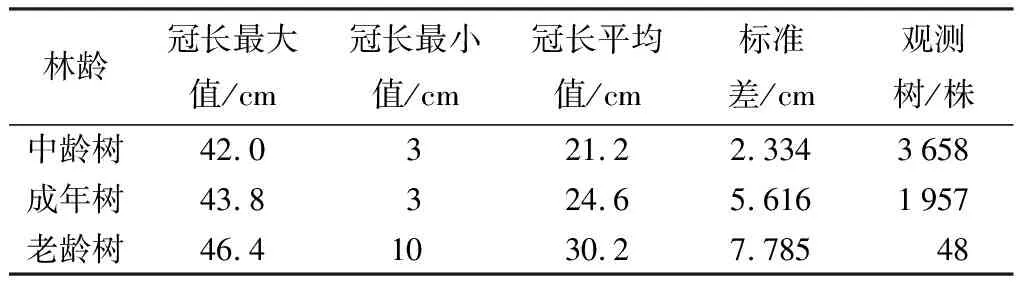
表4 水杉原生种群不同林龄冠长统计
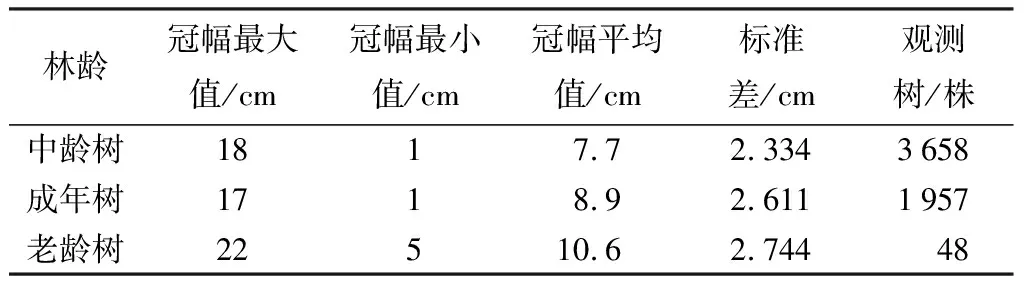
表5 水杉原生种群不同林龄冠幅统计
由表6可知,中龄树的冠幅主要分布在3~9 m,成年树和老龄树的冠幅主要分布在6~12 m,冠幅大于18 m的母树有且仅有1株,冠幅小于3 m的母树仅占中龄树的2.73%,成年树的1.43%。随着年龄增长,3~6 m的冠幅占比大幅度减少,6~9 m的冠幅占比减势微小,几乎持平,9~12 m与12~15 m两级别的冠幅占比呈显著增加。水杉原生种群整体的集中区域从6~9 m往9~12 m偏移。
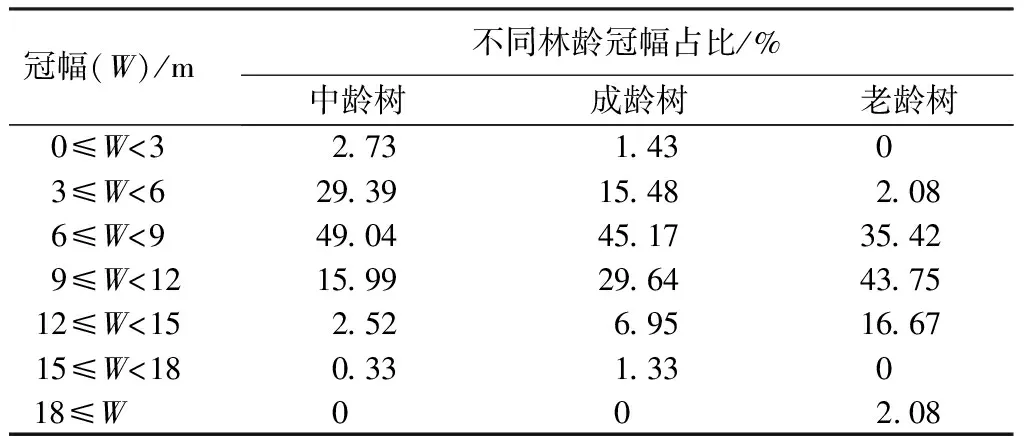
表6 水杉原生种群不同林龄冠幅占比
3.3 冠长率结构
由表7可知,整个原生种群冠长率分布区间为0.148~0.970,中龄树到老龄树变异系数成先减小后增大趋势,说明成年树的冠长率波动较另外两龄级小一些,整体的冠长率呈递增趋势。
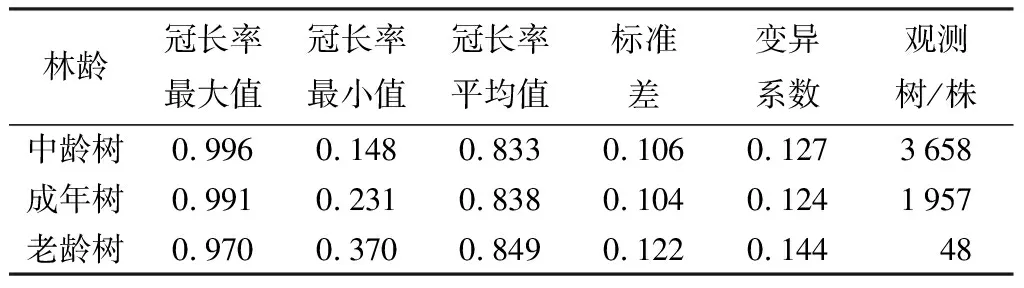
表7 水杉原生种群不同林龄冠长率统计
通过表8可知,中龄树的冠长率主要聚集于0.8~0.9,中龄树的冠长率主要集中于0.8~1.0阶段,老龄树主要分布在0.9~1.0区间,整个水杉原生母树种群冠长率在0~0.1没有分布,随着年龄的增加,0.1~0.3区间的冠长率占比逐渐减少,0.5~0.6、0.7~0.8区间的冠长率占比几乎不变,0.8~0.9区间的冠长率占比呈下降趋势,0.9~1.0区间的冠长率所占比例增加。

表8 水杉原生种群不同林龄冠长率占比
3.4 胸径、树高与树冠因子的通径关系
3.4.1 胸径、树高与冠幅的通径分析
水杉原生种群的胸径、树高和冠幅之间的关系如表9所示。胸径与树高对冠幅的决策系数在不同林龄间都为正数,说明胸径与树高对冠幅都为促进作用。树高与冠幅之间的直接通径系数呈先增大后减小的趋势,且树高对冠幅的正向作用在成年林时最大。胸径对冠幅的正向作用在老龄林中表现显著,不同年龄阶段胸径对冠幅的直接作用均大于间接作用,通过分析直接通径系数和决策系数可知,胸径对于冠幅的影响均大于树高对冠幅的影响。
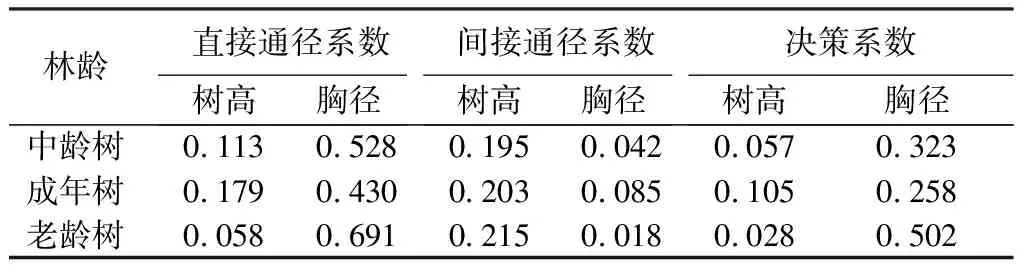
表9 水杉原生种群不同林龄胸径、树高与冠幅的通径分析
3.4.2 胸径、树高与冠长率的通径分析
由表10可知,水杉原生母树在不同林龄阶段树高对冠长率的直接通径系数均大于0,说明树高对冠长率均为正向作用,且在成年林时作用最大。胸径与冠长率之间的直接通径系数整体处于下降的趋势,在成年林达到最小值,表明胸径对于冠高率的正向作用在逐渐减小并一度出现负向作用。

表10 水杉原生种群不同林龄胸径、树高与冠长率的通径分析
3.4.3 胸径树高与冠形率的通径分析
不同林龄水杉原生母树胸径树高和冠形率之间通径分析结果如表11。树高对冠形率的间接作用一直小于直接作用。水杉原生母树处于中龄林时冠形率与胸径和树高之间的直接通径系数均为最大,表明在这个阶段胸径和树高对冠形率有较大的正向作用。树高对冠形率的直接通径系数在成年树达到峰值,整体趋势为呈现先减后增,说明树高对水杉原生母树冠形率的正向作用力先减小后增大。胸径对于冠形率的决策系数呈递减趋势且变为负值,说明胸径对于冠形率正向作用一直减小甚至成为负向作用。
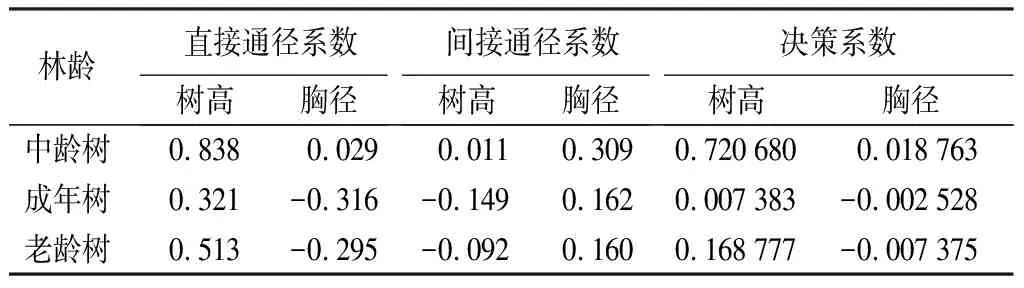
表11 水杉原生种群不同林龄胸径、树高与冠形率的通径分析
4 讨论
4.1 水杉原生种群树冠特征
随着林龄的增加,主要分布冠幅由3~9 m到9~15 m过渡,高级别的冠长率所占比例增加。最大冠幅22 m为谋道1号母树,仅为个例,整个水杉原生种群冠长率在0~0.1没有分布,但0.1~0.3有极少个体分布,这部分个体多为受雷击个体,对比熊彪[4]和黄小[27]对水杉原生种群的研究结果,近二十几年死亡个体中雷击导致的死亡树木约占31.3%,其受雷击影响的个体约占整体1.61%。水杉原生种群中现因雷击和断梢造成个体濒危所占比例均为21.31%,生长状况衰弱的个体中雷击和断梢所占比例为16.49%、30.05%。雷击与断梢降低了树高和冠长,严重影响冠长率,而断梢个体大部分原因也是雷击,所以对水杉原生种群产生主要影响的是雷击。对水杉原生种群的后续保护过程中,可以为较高处于空旷地带的水杉原生母树人为安置避雷设施,例如避雷针等,通过避雷措施使其避免雷害,提高存活率。
通过实地调查结合林勇[5]和Tang et al.[28]对水杉种群的研究,50.89%的水杉个体分布在居民房前屋后,由于水杉个体地处农田、距离房屋太近、前期水杉种子被当地交易当做经济收入来源等原因,水杉个体被人为剃枝,对冠幅宽度、第一活枝高等产生直接影响。部分水杉个体剃枝严重导致生长状况呈濒危状,但仅为个例,对整个水杉原生种群而言影响较小。对水杉原生种群的后续保护过程中,应加强当地居民保护意识,进一步避免人为因素直接导致水杉个体死亡,影响水杉种群树冠结构。
4.2 胸径、树高与树冠因子的关系
通径分析[29]中的直接作用和间接作用能够更加精确地显示各因素的相关关系大小和相对重要性。胸径与树高对冠幅的决策系数在不同林龄间都为正数,说明胸径与树高对冠幅都为促进作用,这与卢妮妮[15]所调查的杉木情况类似。在水杉原生种群不同林龄间,胸径对冠幅的作用呈先减小后增大的趋势,成年树阶段胸径对冠幅作用变小,导致原因可能是受人为影响,早期当地居民人为剃枝采种导致。中龄树和成年树阶段,对冠长率的影响排序为树高大于胸径,在老龄树阶段,对冠长率的影响胸径大于树高,这除了与水杉本身性质有关,还可能是由于部分老龄树遭受雷击断梢导致,所以日后的水杉母树保护中,避免老龄树遭受雷击是重中之重。
树高对冠长率、冠形率均为正向作用,胸径对冠形率冠长率的作用整体呈下降趋势甚至表现为抑制作用。通过比较杉木[15]和水杉,树高对冠长率作用变化趋势相同,成年林阶段胸径对冠长率的间接作用大于直接作用,胸径对冠长率作用在成年林阶段表现为负作用可能是受人为因素影响。通过比较不同林龄间胸径和树高对各冠幅因子的直接作用和间接作用,可以提高水杉母树胸径、树高、冠幅数学模型的精确性,为进一步建立数学模型提供依据。
5 结论
整体来说,水杉原生种群平均冠长和冠幅都在随年龄的变大而增加,且增加趋势近乎一样,冠形率和冠长率都由低级别向高级别过渡,雷击是影响水杉原生种群树冠特征的主要因素。不同林龄阶段的胸径对冠幅的直接作用大于间接作用,它的影响程度大于树高,这与冠形率、冠长率相反。在中龄林和老龄林阶段,胸径对冠长率呈正向作用,整个生长阶段胸径对于冠长率、冠形率之间的直接通径系数整体处于下降的趋势,甚至成熟林均表现为负作用。胸径和树高对冠形率影响在中龄林均为正向作用,在成年林和老龄林,胸径对冠形率呈现显著负向作用,不过胸径对老龄林冠形率的抑制作用小于成年林。通过分析了解到水杉原生种群胸径、树高对冠幅等因素的作用大小,可以结合水杉原生种群生长特性,进一步建立精确的不同林龄阶段的冠幅预测模型,进一步探讨胸径、树高等与天然更新之间的联系,为水杉原生种群的管理和保护提供参考和依据。
猜你喜欢
防护林科技(2022年5期)2022-08-22
现代装饰(2022年3期)2022-07-05
黑龙江气象(2021年2期)2021-11-05
意林·少年版(2021年13期)2021-08-30
数理化解题研究(2021年13期)2021-08-19
机械工程与自动化(2020年6期)2020-12-28
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
中国林副特产(2015年2期)2015-01-25
亚热带资源与环境学报(2015年1期)2015-01-22
