
毛竹竹秆秆柄形态与解剖学研究
2021-11-29 10:20江建平丁雨龙RAMAKRISHNAUMuthusamy
南京林业大学学报(自然科学版) 2021年6期
孙 开,江建平,丁雨龙,RAMAKRISHNAU Muthusamy, 魏 强*
(1.南京林业大学,南方现代林业协同创新中心,江苏 南京 210037; 2.万载县林业局,江西 万载 336100)
毛竹(Phyllostachysedulis)是我国最重要的经济竹种,现有毛竹林面积约443万hm2,占我国人工经营竹林总面积的70%左右[1]。我国竹产业绝大部分的产品以毛竹为原料,作为一种可再生自然资源,其经济价值和生态价值愈来愈受到国内外的广泛关注。近年来,随着高通量转录组测序等新技术在竹学研究中的应用,有关毛竹生长发育基础理论的研究在各方面均获得了长足的进步,如国内外课题组利用高通量转录组测序结合传统发育生物学技术,解析了毛竹地下笋芽的初生增粗生长[2]、竹笋高生长[3-6]、毛竹冷胁迫响应[7]、竹笋采后衰老[8]等重要生物学过程的分子基础。2013年,毛竹基因组测序完成,并在2018年更新到第2版,这大大地促进了毛竹基础理论的研究[9-10]。
竹类结构植物学作为一门传统科学,在帮助竹子研究者揭示竹子生长发育规律、结构与功能方面发挥了重要作用[11]。在过去的几十年间,熊文愈等[12-13]、Liese[14]、丁雨龙等[15-17]一大批竹学研究者对竹秆节间结构、竹秆节部、叶的结构以及根的结构乃至竹秆发育等进行了广泛而详细的研究, 发现了毛竹基本组织具有长、短细胞,节部韧皮部结等特殊细胞与结构,极大地丰富了禾本科植物的解剖学研究理论。
毛竹秆柄是竹秆的一部分,俗称螺丝钉,位于竹秆最基部,与竹鞭相连。相比竹秆其形态极为细小、“节间”显著缩短且不生根,是竹子地上和地下系统连接的重要枢纽[18]。但迄今为止,关于毛竹竹秆秆柄结构与生长发育的研究几近空白。有关竹子竹秆秆柄的研究极少,现有的仅为简单的解剖学观察[19]。本研究从形态学与解剖学两个方面,对毛竹秆柄形态学特征及其解剖学构造进行较为系统的研究,并结合毛竹地下笋芽发育状态,解析其秆柄生长的解剖学与形态学变化,以期丰富竹秆形态建成机制。
1 材料与方法
1.1 试验材料
毛竹各期笋芽及其秆柄于2018年12月采自江西省宜春市万载县大坳村锦源林场(114°21′7.54″E, 28°16′54.58″N),样品采集点土壤类型以红土壤和水稻土为主。每个秆柄剥去芽鳞片之后并做记录,分上、中、下3部分,在体积分数为70%的 FAA[福尔马林-冰醋酸-70%(体积分数)的乙醇]溶液中固定48 h以上待用。 较小的发育前期笋芽(S3期)秆柄剥去芽鳞片之后,保持其与竹鞭连着的状态,固定于体积分数为70%的FAA溶液48 h 以上待用。毛竹地下笋芽发育状态定义参照Wei 等[2],定义如下:①发育前期笋芽(S3期),高约为3.0 cm,芽径约为2.0 cm;②发育中期笋芽(S4期),高约为 5 cm,芽径约为3.5 cm;③发育后期笋芽(S5期),高约为 12 cm,芽径约为 6.0 cm;④成熟笋(S6期),高约为 15 cm,芽径约为8.0 cm。
1.2 形态特征统计
选取毛竹成熟笋秆柄3个,分别记录其附着的芽鳞片数与长度(从秆柄与竹鞭相连部分往上直至第1个长根原基节间以下所有部分)。直径值测定按照秆柄上、中、下3部分分别测定。
1.3 石蜡切片制作与观察
固定好的笋芽秆柄材料经梯度乙醇脱水、二甲苯透明、梯度浸蜡、石蜡包埋后,旋转切片机连续切片,切片厚度为10 μm;切片过程中随机选取部分切片进行镜检以调整切片角度;所得石蜡包埋切片再经体积分数为3%的甲醛溶液展片、37 ℃烘干、二甲苯脱蜡、乙醇脱水、番红-固绿对染、二甲苯透明后,中性树胶封片,用Leica DM2500 (Leica, Wetzlar, Germany)显微镜镜检并拍照。
1.4 滑走切片制作与观察
取上述由FAA固定好的材料使用软化剂(无水乙醇与甘油体积比为1∶1) 软化1个月以上,随后放入聚乙二醇,60 ℃浸泡7 d。自然冷却凝固后,用小刀将已包埋好的聚乙二醇材料块修成小长方体,并将其粘在小木块上。使用滑走切片机(MICROTOME MODEL 860)进行切片,切片厚度为20 μm。随后将切片放入蒸馏水中,洗净聚乙二醇后转入体积分数30%乙醇中脱水20 min,再转入体积分数50%乙醇中继续脱水10 min,之后将切片放入体积分数50%乙醇配制的番红染色液中2 h。取出切片后用体积分数50%乙醇洗掉浮色,再依次转入体积分数分别为70%、80%和95%乙醇中继续梯度脱水,每级10 min。最后浸入体积分数95%乙醇配制的固绿染色液中染色20 s,快速取出后用体积分数95%乙醇洗掉浮色,再依次转入100%乙醇,二甲苯/纯乙醇(体积比1∶1)溶液中,每级10 min。最后在二甲苯中浸泡20 min后,将切片挑至载玻片上,中性树胶封片。待干燥后,利用光学显微镜(Leica DM2500)观察、拍照[20]。
2 结果与分析
2.1 毛竹秆柄基本形态
形态学观察发现,毛竹笋体与竹鞭通过秆柄连接,成熟笋秆柄与芽沟槽大约呈45°角(图1a)。其上附着有芽鳞片,与笋箨类似(图1b)。研究发现,成熟笋秆柄平均长约3.22 cm,上、中、下直径平均分别约为1.91、 1.38 与1.22 cm,附着的芽鳞片数平均约14片(图1d)。
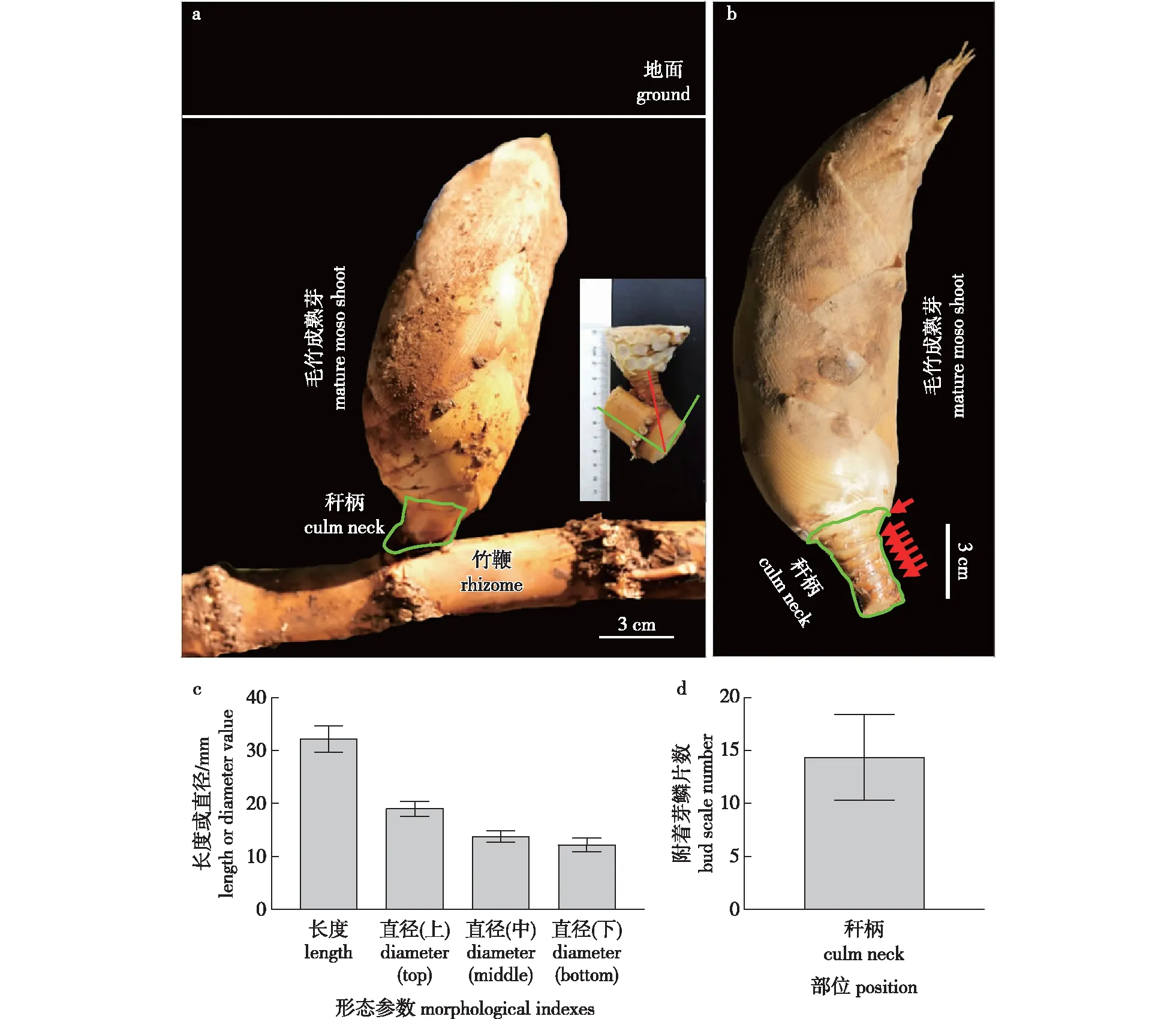
a.连接着笋体与竹鞭的秆柄,右侧小图示秆柄与竹鞭牙槽沟的夹角rhizome and the main body of moso bamboo shoot is connected by a culm neck, the small picture on the right shows the angle between the culm neck and the bamboo rhizome groove position; b. 毛竹成熟笋剥去芽鳞片的秆柄, 红色箭头示芽鳞片附着部位the culm neck of a mature moso bamboo shoot without bud scales, red arrows indicate the positions that the bud scales attach.; c. 毛竹秆柄长度与直径the length and width of moso culm neck; d. 附着在秆柄的芽鳞片数the number of bud scales attached with the culm neck。图1 毛竹秆柄形态Fig.1 Morphology of moso culm neck
2.2 毛竹秆柄解剖学特征
对秆柄中部解剖学观察发现,与竹秆节间中空结构不同,毛竹秆柄为实心结构(图2a)。其横切面布满大量的维管束;从外到内依次分布有表皮、下皮[12]、皮层、维管束及基本组织(图2b、2c)。统计分析发现,下皮平均约有7层细胞,而皮层约有25层细胞,秆柄横切面平均分布有672个维管束(图2d)。
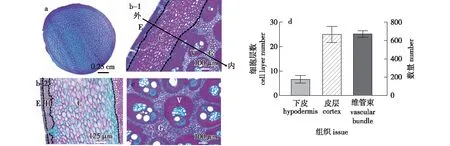
a.毛竹竹秆秆柄横切面 a cross section of moso bamboo culm neck; b. 图a近观图,其中 E、C、V、G 和H 分别表示表皮、皮层、维管束、基本组织和下皮 a close view of figure a. E, C, V, G and H represent epidermis, cortex, vascular bundles, ground tissues and hypodermis, respectively; c. 秆柄维管组织与基本组织vascular tissues and ground tissues of culm neck; d. 毛竹秆柄下皮与皮层细胞层数横切面维管束数量 cell layer of hypodermis and cortex number of vascular bundle in the cross section of culm neck。图2 毛竹秆柄解剖学结构Fig.2 Anatomical structures of moso culm necks
进一步分析发现,毛竹秆柄维管束可初步分为6种类型:①多个维管束纤维帽融合,成长条形分布的维管束,多分布在靠近皮层的地方;②仅具有单个后生导管的维管束(为方便起见,本研究暂定义为单孔维管束);③具有2个后生导管的维管束(本研究暂定义为双孔维管束);④具有3个以上的后生导管的维管束(本研究暂定义为多孔维管束);⑤纤维帽未完全闭合的维管束;⑥纤维帽半开放维管束(图3a)。秆柄维管束形态与竹秆节间和竹鞭节间开放式维管束形态显著不同(图3b、3c)[12,14]。
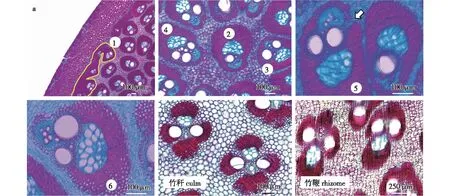
a.数字代表毛竹杆柄维管束类型; 白色箭头示纤维帽未闭合部位 numbers indicate six types of vascular bundles in the culm neck of moso bamboo culm; white arrow indicates the unclosed site of the fiber cap; b. 毛竹竹秆节间典型的维管束形态 typical vascular bundles in the moso bamboo culm internode; c. 毛竹竹鞭节间典型的维管束形态 typical vascular bundles in the moso bamboo rhizome internode。 图3 毛竹维管束类型Fig.3 Vascular bundles in moso bamboo
笔者还发现,毛竹秆柄基本组织细胞纵向排列较为紊乱(图4a),且没有如竹秆节间基本组织的长、短细胞之分(图4b)[14]。

a. 秆柄典型薄壁细胞; 红色闭合线示细胞轮廓 typical parenchyma cells in the moso bamboo culm internode; red enclosed lines represent cell shapes; b.毛竹竹秆薄壁细胞; 红色与绿色闭合线分别代表短细胞与长细胞 parenchyma cells in moso bamboo culm; red and green enclosed lines represent short parenchyma and long parenchyma cells, respectively。图4 毛竹竹秆秆柄与节间薄壁细胞Fig.4 Parenchyma cells in the culm neck and internode of moso bamboo
笔者进一步对成熟笋秆柄从上到下,从外到内的解剖学特征变化进行了观察与分析,根据番红染色深浅程度,初步发现毛竹秆柄底部木质化程度高于中部与上部(图5a、 5b);同时相同放大倍数同一个观察视野下,可观察到秆柄下部的维管束数量要高于中部与上部 (图6)。维管束在秆柄下部以单孔为主逐渐过渡到秆柄上部以双孔为主,而单孔维管束多分布在秆柄中心部位(图6)。

a.毛竹成熟笋秆柄纵切面the vertical section of a culm neck of a mature moso bamboo shoot; b.从上往下秆柄3个部位横切面 three cross sections in different parts along the culm neck from bottom to top。图5 毛竹成熟笋秆柄纵横切面Fig.5 The rertical and cross sections of moso culm neck

秆柄不同部位维管组织vascular tissues in different parts of culm neck; V. 维管组织; 黑色箭头示纤维帽未闭合部位 vascular bundle black arrow indicates the unclosed site vascular bundle of the fiber cap。图6 毛竹竹秆秆柄不同部位维管组织解剖构造Fig.6 Anatomical structures of the moso culm neck in different parts
2.3 不同发育期笋芽秆柄形态与解剖学分析
在上述研究基础上,笔者进一步对不同发育期笋芽秆柄形态与解剖学变化进行了分析(图7)。研究结果表现,与发育前期相比,发育后期笋芽秆柄与成熟笋秆柄长度与粗度接近,秆柄长度显著变长,直径变粗(图7a、7b)。但是,纵切切片显示,在发育前期笋芽秆柄中已观察到15片以上芽鳞片数 (图7c)。
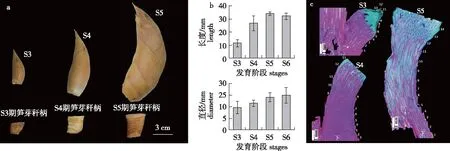
a. 不同发育期毛竹笋芽及其秆柄 bamboo shoots and their culm necks in different stages; b. 不同时期笋芽秆柄长度与直径lengths and diameters of the culm necks in different stages;c. 不同时期笋芽秆柄纵切,数字代表芽鳞片附着位置vertical sections of culm necks in different stages, numbers indicate attached sites of bud scale。图7 不同发育期毛竹笋芽秆柄形态特征与解剖构造Fig.7 Morphologies and anatomical structures of culm necks in moso bamboo shoot at different developmental stages
3 讨 论
毛竹竹秆秆柄是竹秆的一部分,是连接笋体、竹秆与竹鞭的重要桥梁,在竹笋生长过程中是母竹向竹笋运输营养的重要通道。据笔者调查,迄今为止,尚未见有关毛竹竹秆秆柄形态与解剖构造及其生长的报道。
通过传统解剖学结合形态学观察笔者发现,秆柄虽为竹秆的一部分,但其结构与竹秆明显不同[12, 14],如其结构为完全实心,且高度木质化,虽有芽鳞片(与笋箨类似)附着但却无芽着生,因此其没有真正的节。同时其维管束密度高,形态呈现多样,且与竹秆和竹鞭节间开放式维管束形态显著不同[14];其基本组织细胞没有典型的如竹秆节间基本组织有长、短细胞之分[12,14],排列极不规则。与毛竹竹秆下皮细胞层数仅有1~2层的情况相比,其下皮与皮层细胞层数更多[12]。
笔者还对不同发育期毛竹笋芽秆柄的形态与解剖学变化进行了定性分析,发现发育前期笋芽秆柄已具备完整的芽鳞片数(> 15);同时,笋芽秆柄从发育前期至后期具有一个明显的增长与增粗的过程。这些结果暗示发育前期笋芽秆柄基本结构已经分化完成,发育后期笋芽秆柄主要是一个伸长生长的过程。
上述结果表明,毛竹秆柄解剖学构造与竹秆节间等其他部位不同,在今后的研究中,利用高通量转录组测序等最新技术对其展开生长发育动态与基因表达谱研究,解析这个特殊构造形成的生物学机制,对丰富毛竹竹秆形态建成机制具有重要的意义。
猜你喜欢
河北农业大学学报(2021年5期)2021-11-10
湖北林业科技(2021年4期)2021-09-07
林业科学(2020年12期)2021-01-26
新疆农业科学(2020年9期)2020-10-13
创新作文(1-2年级)(2019年8期)2019-12-16
创新作文(小学版)(2019年22期)2019-12-07
河南农业科学(2019年9期)2019-09-24
小学生导刊(2018年4期)2018-04-18
世界竹藤通讯(2017年4期)2017-08-17
小学生导刊(2017年13期)2017-06-15
