
外源半胱氨酸缓解裸燕麦镧胁迫的生理机制
2021-11-22 12:29:28刘建新刘瑞瑞贾海燕刘秀丽卜婷李娜
草业学报 2021年11期
刘建新,刘瑞瑞,贾海燕,刘秀丽,卜婷,李娜
(甘肃省陇东生物资源保护利用与生态修复重点实验室,陇东学院生命科学与技术学院,甘肃庆阳745000)
随着稀土矿的大量开采和稀土的广泛应用,稀土元素通过各种途径进入水体和土壤,对农业生态系统产生深刻影响[1]。稀土微肥在我国应用普遍,有效促进了作物生长发育和逆境耐性增强,并提高产量和改善品质,但长期施用大量稀土微肥导致农田生态系统稀土元素积累,影响作物生长发育,造成减产和品质下降[2]。 镧(lanthanum,La)是稀土肥料中的主要成分之一,也是17 种稀土元素中丰度最大、性质最活泼的元素[3]。植物在遭受La 胁迫时会诱导体内超氧阴离子(O2.−)和过氧化氢(H2O2)等活性氧(reactive oxygen species,ROS)积累,进而引发膜脂过氧化反应,而植物体内的ROS 清除系统在膜脂免受过氧化伤害方面发挥着重要作用[4]。植物在遭受逆境时除ROS 清除系统发生响应外,细胞代谢还会趋向积累渗透溶质来维持细胞膨压,缓解对细胞的不利影响[5]。研究表明,La 胁迫导致黄瓜(Cucumis sativus)细胞膜脂肪酸配比改变[6]、类囊体结构破坏和光系统Ⅱ活性降低[7];烟草(Nicotiana tabacum)碳同化酶活性下降[8];裸燕麦(Avena nuda)和黑麦草(Lolium perenne)矿质元素平衡破坏、碳氮代谢失调和光合作用受抑[4,9−10];番茄(Lycopersicon esculentum)干物质积累下降[11]。可见La 胁迫诱导细胞代谢紊乱和生长受抑等胁迫伤害。因此,探寻缓解作物La 胁迫伤害的技术途径,对克服稀土应用带来的负面效应具有重要实际意义。
硫化氢(hydrogen sulfide,H2S)是植物体内新发现的第3 种气体信号分子,它参与植物逆境响应调节,能够缓解重金属对植物产生的胁迫伤害[12]。植物体内的H2S 可通过多条途径产生,目前认为半胱氨酸(cysteine,Cys)通过Cys 脱巯基酶催化产生H2S 可能是主要途径[13]。L-半胱氨酸(L-cysteine,L-Cys)是组成蛋白质的20 种基本氨基酸之一,也是植物无机硫酸盐同化为有机硫化物途径中的终产物及参与硫素代谢的中间载体。Cys 的巯基可与重金属盐络合形成硫醇盐,也可与有毒芳香族化合物缩合成硫醚氨酸而解毒;由Cys、谷氨酸和甘氨酸合成的谷胱甘肽(glutathione,GSH)以及由GSH 为前体合成的植物络合素(phytochelatins,PCs)在植物抗氧化和螯合重金属方面发挥着重要作用[14];Cys 与胱氨酸相互转换对稳定蛋白质空间结构和保持其活性也具有重要影响[15]。有研究表明,外源Cys 可延缓乙烯诱导的橡胶(Hevea brasiliensis)叶片衰老[16];缓解SO2对大麦(Hordeum vulgare)的氧化损伤[17];增强小麦(Triticum aestivum)对铜胁迫的耐性[18]。而外源Cys 能否缓解植物La 胁迫伤害,迄今的研究尚未见报道。裸燕麦是我国北方广泛种植的一种高蛋白、富含膳食纤维等营养和保健成分的禾本科燕麦属杂粮作物[19]。北方城市内蒙古包头市的稀土储量约占中国储量的85%,长达60 余年的稀土矿开采使矿区土壤稀土元素含量超过内蒙古几何背景值的481 倍[20],白云鄂博矿区0~60 cm 表层土壤La 含量达到107. 15~2031. 7 mg·kg−1[21]。裸燕麦具有较强抗逆性,在稀土污染等质地差的边际土壤常首选种植裸燕麦,故裸燕麦的生长发育遭受La 胁迫伤害。本试验通过盆栽土培试验,研究喷施不同浓度L-Cys 对La 胁迫下裸燕麦生长的影响及生理响应的调节效应,探讨外源Cys 对裸燕麦La 胁迫的缓解作用及其生理机制,以期为利用Cys 缓解作物La 胁迫伤害提供理论依据。
1 材料与方法
1. 1 供试材料
试验于2018 和2019 年的3−7 月在甘肃省陇东生物资源保护利用与生态修复重点实验室试验基地进行。供试土壤取自陇东黄土高原董志塬(35°36′47″N,107°34′44″E)黑垆土0~20 cm 耕层土壤,土壤有机质10. 84 g·kg−1,全氮0. 744 g·kg−1,全磷(P2O5)0. 582 g·kg−1,全钾(K2O)17. 3 g·kg−1,碱解氮35. 2 mg·kg−1,速效磷(P2O5)4. 52 mg·kg−1,速效钾(K2O)146. 9 mg·kg−1,pH 8. 4,土壤La 含量为0. 059 mg·kg−1。供试裸燕麦品种为‘定莜9号’,种子由甘肃省定西市农业科学研究院提供。供试L-Cys 为Sigma-Aldrich 公司产品,La 供体氯化镧(LaCl3·2. 5H2O)购自上海生物工程技术服务有限公司。
1. 2 试验设计
1. 2. 1 La 胁迫下裸燕麦生长试验 称取氯化镧和过5 mm 筛风干土,拌匀,使土壤La 含量分别为25,50,100,200,300,400,500 mg·kg−1(含土壤La 背景值),以不添加氯化镧为对照(CK)。分别称取20. 0 kg 拌匀土样与10. 0 g 复合肥(N∶P2O5∶K2O=13∶17∶15)混匀,装入口径31. 5 cm、高34. 0 cm 的塑料盆钵中,按田间持水量(75±5)%浇清水平衡1 周后进行播种(2018 年3 月28 日),每盆播裸燕麦种子约80 粒,播种深度5 cm。2018 年4 月24日间苗,5 月2 日疏苗,每盆保留一致壮苗25 株左右。5 月20 日和6 月9 日各追肥1 次,叶面喷施1. 0% 尿素和0. 5% 磷酸二氢钾每盆50 mL,常规管理。7 月15 日成熟收获,测量株高、地上部生物量、穗粒数、穗粒重、千粒重和籽粒产量。每处理1 盆,重复5 次,随机排列。
1. 2. 2 外源Cys 缓解裸燕麦La 胁迫试验 参考文献[18]中L-Cys 喷施的有效浓度0. 5~5. 0 mmol·L−1。本盆栽土培试验设3 组,共6 个处理:1)对照组(CK):盆栽土壤不施外源La,植株叶面喷施蒸馏水;2)La 胁迫组:添加氯化镧使盆栽土壤含La 400 mg·kg−1(含土壤La 背景值),植株叶面喷施蒸馏水;3)Cys 处理组:添加氯化镧使盆栽土壤含La 400 mg·kg−1(含土壤La 背景值),并分别在植株叶面喷施0. 5、1. 0、2. 5、5. 0 mmol·L−1L-Cys 溶液。20. 0 kg 过5 mm 筛风干黑垆土,拌入氯化镧(CK 组不添加氯化镧)和10. 0 g 复合肥(N∶P2O5∶K2O=13∶17∶15),混匀,装入塑料盆钵(口径31. 5 cm、高34. 0 cm)中,每盆浇9 L 水后平衡7 d 播种裸燕麦(2019 年3 月25 日),每盆播80 粒左右种子,播后覆5 cm 土。4 月8 日出苗后根据生长情况及时间苗和疏苗,每盆定苗25 株。定期追肥,常规管理。在裸燕麦抽穗期(2019 年6 月12 日)给Cys 处理组叶面喷施不同浓度Cys 溶液,向CK 组和La 胁迫组喷施等量蒸馏水。喷施早晚各喷一次,连喷3 d,每次喷至正反面叶片溶液欲滴为止,喷施量每盆约40 mL。为提高喷施液与叶面的粘附程度,喷施液含0. 01% 表面活性剂吐温−80(迈瑞尔化学技术有限公司,上海)。每处理4 盆,5 次重复,随机排列。最后1 次喷施后的第7 和14 天分别取倒2 叶用液氮速冻30 s 后超低温冰箱−70 ℃保存,测定相关指标。成熟后测定根系、茎秆和叶片La 含量,收获(2019 年7 月10 日)并进行室内考种,调查株高、地上部生物量、穗粒数、穗粒重、千粒重和籽粒产量。
1. 3 测定指标与方法
采用羟胺反应α-萘胺显色法测定O2·−含量,硫代巴比妥酸比色法测定丙二醛(malondialdehyde,MDA)含量[22];按Sergiev 等[23]的方法测定H2O2含量;参照陈建勋等[24]的方法测定超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POD)和抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性及抗坏血酸(ascorbic acid,ASA)和GSH 含量。类胡萝卜素含量采用乙醇提取比色法测定[22];PCs 含量测定参照白晓娟等[25]的方法;可溶性蛋白质、可溶性糖和脯氨酸含量用高俊凤[22]的方法测定。采用Prodigy 型电感耦合等离子体发射光谱仪(Lee man 公司,美国)分别测定根系、茎秆和叶片中La 含量[4],并计算对La 的富集系数和转运系数。富集系数=裸燕麦根系、茎秆或叶片La 含量/土壤La 含量,式中:土壤La 含量=试验中添加的La 总量(含土壤背景值)/盆土质量。转运系数=裸燕麦库组织La 含量/裸燕麦源组织La 含量,La的转运系数划分为从根向地上部的转运系数、根向茎的转运系数和茎向叶的转运系数。
1. 4 数据统计与分析
所有数据采用Micorsoft Excel 2007 计算整理和制图,SPSS 20. 0 软件进行SSR 法差异显著性分析(P<0. 05),结果以平均值±标准差表示。
2 结果与分析
2. 1 La 胁迫对裸燕麦生长和产量构成因素的影响
由表1 可见,除25 mg·kg−1La 处理略微提高裸燕麦株高、地上部生物量、穗粒数和籽粒产量外,随着La 含量增加,株高、地上部生物量、穗粒数、穗粒重和籽粒产量显著降低,而千粒重变化不大。与CK 相比,≤100 mg·kg−1La 处理的株高、穗粒重差异不显著,≤200 mg·kg−1La 处理的地上部生物量、穗粒数和籽粒产量差异无统计学意义,≥300 mg·kg−1La 处理显著抑制裸燕麦的生长及籽粒产量和产量因素构成。经回归分析,裸燕麦籽粒产量(y)与La 含量(x)极显著负相关:y=29. 164−0. 037x(R2=0. 908**),裸燕麦籽粒产量降低1/2 时的La 含量(EC50)=404. 97 mg·kg−1。因此,研究外源Cys 对裸燕麦La 胁迫缓解作用时使用La 含量400 mg·kg−1。

表1 外源La 对裸燕麦株高、地上部生物量和产量构成因素的影响Table 1 Effect of exogenous lanthanum on plant height,shoot biomass and yield component factors of naked oat
2. 2 外源Cys 对La 胁迫下裸燕麦叶片O2. −、H2O2和MDA 含量的影响
由表2 可知,与CK 相比,La 胁迫提高了裸燕麦叶片O2.−、H2O2和MDA 含量,三者含量随胁迫时间延长进一步提高(P<0. 05)。La 胁迫下,喷施0. 5 和1. 0 mmol·L−1Cys 后第7 和14 天裸燕麦叶片中的O2·−、H2O2和MDA含量增幅显著得到缓解,且喷施1. 0 mmol·L−1Cys 的缓解作用大于喷施0. 5 mmol·L−1Cys。La 胁迫下喷施2. 5和5. 0 mmol·L−1Cys 后第7 天的O2·−含量与单一La 处理差异不显著,H2O2和MDA 含量显著降低;喷施2. 5 mmol·L−1Cys 后第14 天的O2·−、H2O2和MDA 含量显著低于单独La 处理,而喷施5. 0 mmol·L−1Cys 的O2·−和H2O2含量差异不显著,MDA 含量显著下降。
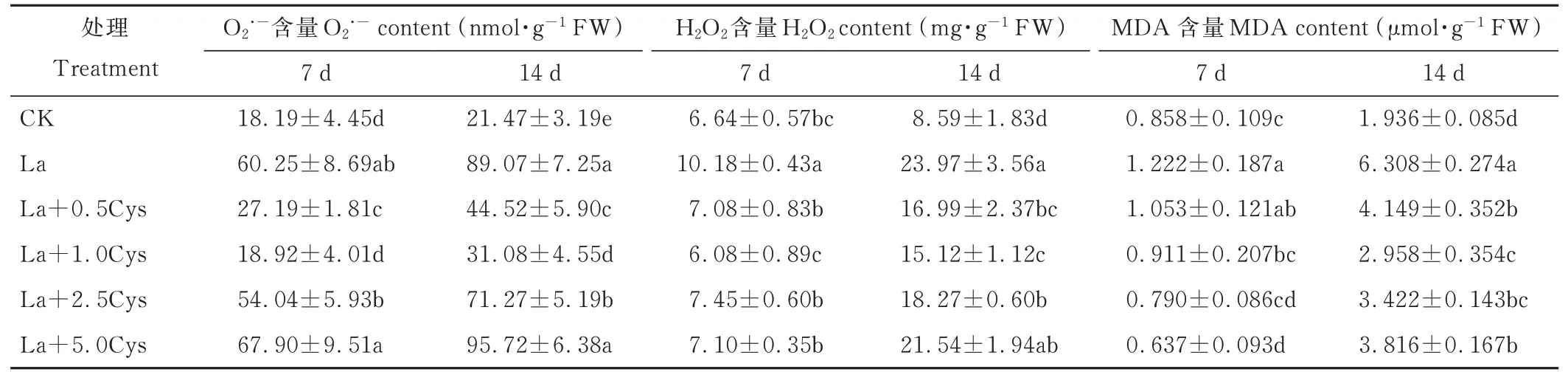
表2 外源Cys 对La 胁迫下裸燕麦叶片O2.-、H2O2和MDA 含量的影响Table 2 Effect of exogenous Cys on O2.-,H2O2 and MDA contents in leaves of naked oat under La stress
2. 3 外源Cys 对La 胁迫下裸燕麦叶片抗氧化酶活性的影响
与CK 相比,La 胁迫显著提高了裸燕麦叶片SOD、CAT 和POD 活性,显著降低APX 活性(图1)。与单一La处理相比,喷施不同浓度Cys 均能降低La 胁迫下SOD 活性,其中2. 5 mmol·L−1Cys 的SOD 活性最低;CAT 活性在Cys 浓度为0. 5 和1. 0 mmol·L−1时显著升高,2. 5 和5. 0 mmol·L−1时CAT 活性降低;喷施0. 5 和2. 5 mmol·L−1Cys 第7 天的POD 活性与单一La 处理无显著差异,喷施1. 0 和5. 0 mmol·L−1Cys 第7 天的POD 活性低于单一La 处理,喷施不同浓度Cys 第14 天的POD 活性均显著低于单一La 处理;La 胁迫下喷施不同浓度Cys 均显著提高了APX 活性,喷施2. 5 mmol·L−1Cys 的APX 活性最大。

图1 外源Cys 对镧胁迫下裸燕麦叶片SOD、CAT、POD 和APX 活性的影响Fig. 1 Effect of exogenous Cys on the activities of SOD,CAT,POD and APX in naked oat leaves under La stress
2. 4 外源Cys 对La 胁迫下裸燕麦叶片抗氧化物质和PCs 含量的影响
La 处理裸燕麦叶片的ASA、GSH、类胡萝卜素和PCs 含量均显著高于CK(图2)。La 胁迫下喷施不同浓度Cys 后ASA 和类胡萝卜素含量明显降低,其中2. 5 mmol·L−1Cys 处理的ASA 含量最低,不同浓度Cys 处理间的类胡萝卜素含量无显著差异;喷施Cys 显著提高了La 胁迫下的GSH 和PCs 含量,并随Cys 浓度增加GSH 和PCs含量总体呈上升的趋势。
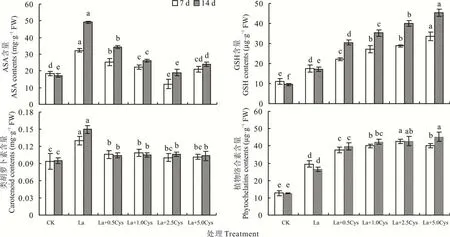
图2 外源Cys 对镧胁迫下裸燕麦叶片ASA、GSH、类胡萝卜素和PCs 含量的影响Fig. 2 Effect of exogenous Cys on the contents of ASA,GSH,carotenoid and phytochelatins in naked oat leaves under La stress
2. 5 外源Cys 对La 胁迫下裸燕麦叶片可溶性蛋白质、可溶性糖和脯氨酸含量的影响
La 处理下裸燕麦叶片可溶性蛋白质含量与CK 差异不显著(7 d)或显著下降(14 d),喷施0. 5~5. 0 mmol·L−1Cys 不同程度提高了La 胁迫下的可溶性蛋白质含量,但不同浓度Cys 处理间的可溶性蛋白质含量差异不显著(表3)。与CK 相比,La 处理的可溶性糖含量显著降低,脯氨酸含量显著增加;La 胁迫下喷施0. 5~5. 0 mmol·L−1Cys 不同程度提高了可溶性糖和脯氨酸含量,其中2. 5 mmol·L−1Cys 处理的增幅最大。

表3 外源Cys 对镧胁迫下裸燕麦叶片可溶性蛋白质、可溶性糖和脯氨酸含量的影响Table 3 Effect of exogenous Cys on contents of soluble protein,soluble sugar and proline in naked oat leaves under La stress(mg·g-1 FW)
2. 6 外源Cys 对La 胁迫下裸燕麦生长和产量构成因素的影响
La 胁迫显著降低了裸燕麦株高、地上部生物量及穗粒数、穗粒重和籽粒产量,分别比CK 下降了15. 6%、40. 3%、42. 2%、51. 0% 和47. 5%,而千粒重无显著变化(表4)。La 胁迫下喷施不同浓度Cys 对裸燕麦株高和千粒重的影响不显著,而对地上部生物量、穗粒数、穗粒重和籽粒产量均有不同程度的提升作用,其中对地上部生物量、穗粒数和籽粒产量作用最大的Cys 浓度是2. 5 mmol·L−1,对穗粒重作用最大的Cys 浓度是1. 0 mmol·L−1。

表4 外源Cys 对镧胁迫下裸燕麦生长和产量构成因素的影响Table 4 Effect of exogenous Cys on the growth and yield component factors in naked oat under La stress
2. 7 外源Cys 对La 胁迫下裸燕麦La 吸收积累特征的影响
La 胁迫显著提高了裸燕麦根、茎、叶对La 的吸收积累量,降低了对La 的富集能力,吸收积累量和富集系数根系>茎秆>叶片;随Cys 浓度增大La 胁迫下裸燕麦根、茎、叶对La 吸收积累量显著增加,富集系数也呈增加趋势(图3)。La 胁迫下根系向地上部和茎向叶的转运系数显著提高,根向茎的转运系数变化不明显;La 胁迫下除喷施0. 5 mmol·L−1Cys 使根系向地上部和茎向叶的转运系数显著下降外,喷施其他浓度Cys 的根系向地上部和茎向叶的转运系数及喷施所有浓度Cys 的根系向茎的转运系数无显著变化。

图3 外源Cys 对La 胁迫下裸燕麦La 含量及富集系数和转运系数的影响Fig. 3 Effect of exogenous Cys on the lanthanum content,accumulator factor and translocation factor in naked oat under La stress
3 讨论
3. 1 外源Cys 对La 胁迫下裸燕麦La 吸收积累和生长特性的影响
研究表明,400 mg·kg−1La 胁迫使裸燕麦对La 吸收积累显著增加,且吸收积累量根系>茎秆>叶片,而根系、茎秆和叶片对La 的富集系数大幅下降,根向地上部、根向茎和茎向叶的转运系数不同程度增大(图3)。说明La 胁迫下裸燕麦各部位对La 的富集能力降低,但各部位间的转运能力和吸收积累量明显提高。 这与前人在牧草植物圆叶决明(Chamaecrista rotundifolia)上的研究结果类似[26]。 在抽穗期喷施0. 5~5. 0 mmol·L−1Cys 显著促进了400 mg·kg−1La胁迫下裸燕麦根系、茎秆和叶片对La 的吸收积累,提高了根、茎和叶对La 的富集能力,且吸收积累量和富集系数随Cys 浓度增加而提高,而对裸燕麦各部位间La 的转运系数几乎无影响。说明外施Cys 可增强裸燕麦对La 的富集能力,促进La 的吸收积累,但对La的转运影响不大。这可能是Cys 作为硫代谢的中间载体和GSH 的合成原料,合成GSH 后进而参与PCs合成[14],PCs 与进入体内的La3+形成PCs−La3+复合体进而转运至液泡,从而促进La3+富集和吸收积累[27]所致。
研究已证明,La 对植物生长的影响表现为“低促高抑”,但不同植物耐受La 的能力存在差异[28−29]。本研究显示,25 mg·kg−1土壤La 不同程度提高了裸燕麦株高、地上部生物量、穗粒数和籽粒产量;但随着La 含量进一步提高,裸燕麦株高、地上部生物量、穗粒数、穗粒重和籽粒产量明显下降,而千粒重变化不大(表1)。说明高浓度La抑制裸燕麦生长和产量构成。这与红壤中La 积累抑制水稻(Oryza sativa)生长发育和产量的结果一致[30]。在400 mg·kg−1La 胁迫下,喷施0. 5~5. 0 mmol·L−1Cys 对裸燕麦株高和千粒重没有影响,但不同程度提高了地上部生物量、穗粒数、穗粒重和籽粒产量,其中2. 5 mmol·L−1Cys 提升地上部生物量和籽粒产量的作用最大(表4)。说明喷施适宜浓度Cys 可以缓解La 胁迫对裸燕麦生长及产量构成的抑制。这与彭向永等[18]研究的外源Cys 能够缓解铜胁迫对小麦幼苗生长抑制的结果类似。
3. 2 外源Cys 对La 胁迫下裸燕麦抗氧化系统和渗透胁迫的调节作用
试验表明,400 mg·kg−1La 胁迫使裸燕麦叶片O2.−、H2O2和MDA 含量提高(表2),可溶性糖和可溶性蛋白质含量下降(表3)。说明La 胁迫诱导ROS 积累造成裸燕麦生理代谢产生紊乱。ROS 积累一方面可造成细胞氧化损伤,同时也可诱导植物抗氧化机制建立[10]。本试验中,La 胁迫使裸燕麦叶片SOD、CAT 和POD 活性(图1)及ASA、GSH、PCs、类胡萝卜素(图2)和脯氨酸(表3)含量提高,但APX 活性降低。说明裸燕麦通过调控抗氧化系统增强适应La 胁迫的能力。可能因为APX 活性下降还是导致ROS 产生积累,造成细胞氧化伤害。植物可以Cys 为前体合成GSH、PCs 和金属硫蛋白等在抵抗重金属胁迫中发挥重要作用[14]。La 这种轻稀土元素本身也是一种重金属元素。本试验中,喷施0. 5~2. 5 mmol·L−1Cys 降低了400 mg·kg−1La 胁迫下裸燕麦叶片O2·−含量,喷施0. 5~5. 0 mmol·L−1Cys 降低了H2O2和MDA 含量(表2)。说明施用Cys 可以降低裸燕麦La 胁迫诱导的氧化损伤。Cys 通过Cys 脱巯基酶催化可产生H2S[13]。H2S 作为第3 种气体信号,能够缓解植物重金属胁迫伤害[12]。外施Cys 对裸燕麦La 胁迫氧化损伤的缓解作用是否与Cys 作为H2S 供体促进H2S 产生而发挥作用有关,尚需进一步探究。La 胁迫下喷施0. 5~5. 0 mmol·L−1Cys 降低了裸燕麦叶片SOD 和POD 活性,提高了APX 活性;喷施0. 5 和1. 0 mmol·L−1Cys 使La 胁迫下裸燕麦叶片CAT 活性增强,喷施2. 5 和5. 0 mmol·L−1Cys 时CAT 活性却显著下降(图1)。另外,喷施0. 5~5. 0 mmol·L−1Cys 降低了La 胁迫下裸燕麦叶片ASA 和类胡萝卜素含量,提高了GSH 和PCs 含量(图2)。这说明外源Cys 能够通过调控抗氧化酶活性和抗氧化物质含量降低La 胁迫诱导的ROS 积累对裸燕麦造成的氧化伤害。这与Deng 等[31]的N-乙酰-L-半胱氨酸可通过增加GSH 和蛋白巯基保护龙葵(Solanum nigrum)免受镉胁迫氧化应激损伤的研究结果类似。
La 胁迫扰乱细胞碳氮代谢,使之趋向积累渗透溶质方向转变[10]。可溶性糖、可溶性蛋白质和脯氨酸等是重要的有机渗透调节剂,其含量增加对维持逆境下植物细胞膨压具有重要作用,其中可溶性蛋白质还具有缓解重金属毒害和保护生物膜的作用[32]。脯氨酸又可作为金属螯合剂和蛋白质稳定剂,还参与细胞ROS 清除[33]。本试验中,喷施0. 5~5. 0 mmol·L−1Cys 可缓解400 mg·kg−1La 胁迫下裸燕麦叶片可溶性蛋白质和可溶性糖含量的下降,进一步提高脯氨酸含量,其中2. 5 mmol·L−1Cys 的效果最佳(表3)。说明施用Cys 增强渗透调节可能也是其缓解La 胁迫伤害的重要机制之一。但Cys 促进La 胁迫下渗透调节物质积累的具体机制有待进一步探讨。
4 结论
喷施适宜浓度Cys 可缓解La 胁迫对裸燕麦生长及产量构成因素的抑制,降低O2.−、H2O2、MDA、ASA、类胡萝卜素含量和SOD、POD 活性,提高CAT、APX 活性及GSH、PCs、可溶性蛋白质、可溶性糖和脯氨酸含量,其最有效的Cys 浓度为1. 0 和2. 5 mmol·L−1。另外,喷施Cys 还可促进La 胁迫下裸燕麦根、茎和叶对La 的富集和吸收积累,但对裸燕麦株高、千粒重及La 在不同部位的转运影响不大。表明外源Cys 能够通过调控抗氧化系统和促进渗透溶质积累以及改变La 的吸收积累,减轻La 胁迫诱导的氧化伤害和对农艺产量性状的抑制。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:39:40
舰船科学技术(2022年11期)2022-07-15 07:51:56
现代畜牧科技(2021年4期)2021-12-05 15:34:45
现代畜牧科技(2021年10期)2021-11-19 08:42:26
西藏农业科技(2019年3期)2019-11-04 00:35:10
新农村(2018年2期)2018-08-18 01:42:28
河北农业科学(2018年2期)2018-07-26 11:28:14
现代园艺(2018年3期)2018-02-10 05:18:12
商洛学院学报(2017年2期)2017-05-17 05:19:53
上海农业学报(2017年3期)2017-04-10 12:39:12
