
镰刀菌根腐病拮抗菌的筛选及鉴定
2021-11-19 07:35樊炳君姚丽段娇罗昆艳杨雪刚焦钰朱国兴魏薇曹艳茹
江苏农业科学 2021年20期
樊炳君 姚丽 段娇 罗昆艳 杨雪刚 焦钰 朱国兴 魏薇 曹艳茹


摘要:为了获得具有抑制镰刀菌根腐病的生防菌,从研究较少的兰坪铅锌尾矿极端环境中采集样品分离菌株,运用平板对峙法从中进行4种镰刀菌根腐病拮抗菌的初筛,共获得10株有抑制效果的菌株。通过平板对峙法和发酵液涂布法进行复筛,获得1株抑菌活性较强的拮抗菌株KC 121。根据生理生化特性和16S rDNA测序结果分析,将KC 121菌株鉴定为特基拉芽孢杆菌(Bacillus tequilensis)。抑菌试验结果显示,KC 121对根腐病的4个病原菌(腐皮镰刀菌、禾谷镰刀菌、尖孢镰刀菌和拟枝孢镰孢菌)均有不同程度的抑制效果。同发酵液相比,菌体的抑菌效果较好。KC 121对拟枝孢镰孢菌的抑制效果达82.66%,对腐皮镰刀菌的抑菌率达76.91%。KC 121的发酵液对拟枝孢镰孢菌和腐皮镰刀菌的抑菌率分别为84.76%和60.85%。本试验结果可以为根腐病的生物防治提供菌种资源,具有进一步的研究和开发价值。
关键词:镰刀菌属;根腐病;生防菌;鉴定;特基拉芽孢杆菌
中图分类号: S432.4+4 文献标志码: A
文章编号:1002-1302(2021)20-0132-06
收稿日期:2021-02-27
基金项目:国家自然科学基金(编号:31660002);云南省教育厅科学研究基金(编号:2020Y0475);云南省地方本科高校(部分)基础研究联合专项资金(编号:2018FH001-003);云南省“万人计划”青年拔尖人才资助项目(编号:YNWR-QNBJ-2018-011);省部共建云南生物资源保护与利用国家重点实验室开放课题(编号:2019KF005)。
作者简介:樊炳君(1996—),女,云南昆明人,硕士研究生,主要从事资源利用与植物保护研究。E-mail:fanbingjun928@sina.com。
通信作者:曹艳茹,博士,副教授,主要从事微生物多样性及功能挖掘研究。E-mail:yanrucao3@aliyun.com。
镰刀菌(Fusarium)隶属于菌物界真菌门,有性型是子囊菌门肉座菌的赤霉属(Gibberella)、丛赤壳属(Nectria)、丽赤壳属(Calonectria)和小赤壳属(Micronectriella),无性型是半知菌亚门瘤座孢菌科[1]。由于该属菌株着生于子座上且具有月牙形或船形的分生孢子,1908年Link首次将其命名为镰刀菌,并以粉红镰刀菌为模式种建立了镰刀菌属[2]。随着科学技术的不断进步,镰刀菌属的分类标准也逐步完善,目前被广泛接受的种类约70余种[1]。作为重要的植物病原菌,镰刀菌属的菌株可危害众多粮食作物、蔬菜、水果、中药材和花卉等,造成严重的经济损失[3-6]。镰刀菌不仅会使作物萎蔫、腐烂和坏死,产生的毒素被人畜食用后也会引发中毒[7]。因此,针对该属的防治研究一直受到人们的关注和重视。
镰刀菌是一种土壤习居菌,以菌丝和孢子的方式越冬,常分布在植株的病残体、带病的种子和条件适宜的土壤中,具有寄主种类多、致病能力强和危害严重等特点[8]。目前对于镰刀菌的防治措施主要是化学防治,应用于实际生产的生物防治研究还相对较少。刘利佳等通过筛选发现,62.5 g/L的精甲·咯菌腈悬浮种衣剂对尖孢镰刀菌(Fusarium oxysporium)的抑菌率达97.27%,是防治烟草镰刀菌根腐病效果最佳的化学药剂[9]。王国荣等研究发现甲基硫菌灵和戊唑醇及其复配制剂对芹菜茎基腐病病原菌尖孢镰刀菌的抑制效果最好;在田间药效试验中发现,75%肟菌·戊唑水分散粒剂(WDG)和70%甲基硫菌灵可湿性粉剂(WP)的田间防效较好[10]。肖荣凤等通过筛选发现2 000倍液的6.6%嘧菌酯·1.1%咯菌腈·3.3%精甲霜灵悬浮种衣剂和25%吡唑醚菌酯乳油对太子参根腐病病原菌尖孢镰刀菌的抑菌率均高于75%[11]。许乐等从丹参植株中分离获得一株内生细菌多粘类芽孢杆菌(Paenibacillus polymyxa)能够有效抑制丹参根腐病病原菌腐皮镰刀菌(Fusarium solani)的生长[12]。牛世全等从盐碱土壤中分离获得一株枯草芽孢杆菌(Bacillus subtilis)对黄芪根腐病病原菌尖孢镰刀菌的抑制效果较好[13]。
运用生态环保的生物菌剂防治镰刀菌属菌株引起的病害,在绿色可持续防治病害的同时,还能保护人畜的健康。本研究从兰坪铅锌尾矿极端环境中分离原核微生物,从中筛选镰刀菌根腐病的拮抗菌,為生物菌剂的研发提供更多菌种资源。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 分离自笔者所在课题组于2019年9月采集的兰坪铅锌尾矿样品,采样地的海拔范围在 2 208~2 825 m,经纬度范围为26°22′~26°24′N、99°23′~99°28′E,年平均气温为10.4~11.8 ℃,属温带山地主体型季风气候[14]。将重复菌株去除后,共从24个样品中分离得到83个种的菌株,隶属于4个门6个纲16个目22个科30个属,具体信息见表1。
1.1.2 病原菌 腐皮镰刀菌Fusarium solani(编号B6)、禾谷镰刀菌Fusarium graminearum(编号B7)和尖孢镰刀菌Fusarium oxysporum(编号B8)购买自中国农业微生物菌种保藏管理中心;拟枝孢镰刀菌Fusarium sporotrichioides(编号B9)购买自北京北纳创联生物技术研究院。
1.2 菌株培养
1.2.1 供试菌株培养 从保藏试管中挑取供试菌株接种在ISP 2培养基上,置于28 ℃恒温箱中培养 3~7 d[15]。
1.2.2 病原菌培养 从保藏试管中挑取病原菌接种在马铃薯葡萄糖琼脂(PDA)培养基上,置于 25 ℃ 恒温箱中培养7 d[16]。
1.3 拮抗菌的初筛
运用平板对峙法[17]将供试菌株接种于ISP 2平板上(10株/皿),于28 ℃恒温箱中培养5~7 d后,在平板中央接种直径为8 mm的病原菌菌块,在 25 ℃ 恒温箱中培养7 d后观察抑菌效果,对有抑菌活性的阳性菌株进行复筛。
1.4 拮抗菌的复筛
1.4.1 供试菌体对病原菌生长的影响 运用平板对峙法把筛选出的阳性菌株按4点法接种于PDA平板上,方法同“1.3”节,每个处理重复3组,以不接供试菌株为空白对照。
1.4.2 供试菌株发酵液对病原菌生长的影响 把筛选出的阳性菌株接种于ISP 2液体培养基,置于200 r/min 28 ℃的摇床培养5 d后,离心收集上清液,得到菌株的发酵液,再将其用0.22 μm的细菌过滤器进行过滤。将200 μL过滤发酵液涂布于PDA平板,待培养基表面将液体吸收后将直径为 8 mm 的病原菌菌块接种于其上[18],在25 ℃恒温箱培养 7 d 后观察抑制效果并计算抑菌率[19],每个处理重复3组,以清水涂板为空白对照。所获数据通过SPSS Statistics软件进行差异显著性分析。
抑菌率=对照组菌落直径-处理组菌落直径对照组菌落直径-0.8 cm×100%。
1.5 拮抗菌的鉴定
1.5.1 形态观察 在ISP 2培养基上将“1.4”节中筛选出的阳性菌株进行划线培养,28 ℃培养3 d后观察菌落生长情况及形态特征。
1.5.2 生理生化特性检验[20] 测定菌株产生接触酶、脲酶和酯酶的能力,检测菌株水解淀粉、明胶液化、牛奶胨化、分解纤维素、还原硝酸盐和产生硫化氢的能力,同时进行甲基红(MR)试验和V-P试验。
1.5.3 分子鉴定 采用酶法小量DNA提取法提取菌体的基因组DNA[15],运用16S rRNA基因的通用引物(PA:5′-CAGAGTTTGATCCTGGCT-3′和PB:5′-AGGAGGTGATCCAGCCGCA-3′)[21]进行PCR特异扩增,随后送到生工生物工程(上海)股份有限公司进行测序。将获得的16S rRNA基因序列提交到EzBioCloud(https://www.ezbiocloud.net/identify)进行比对,用MEGA 7软件采用邻接法(Neighbor-Joining)构建系统发育树,确定菌株种属。
2 结果与分析
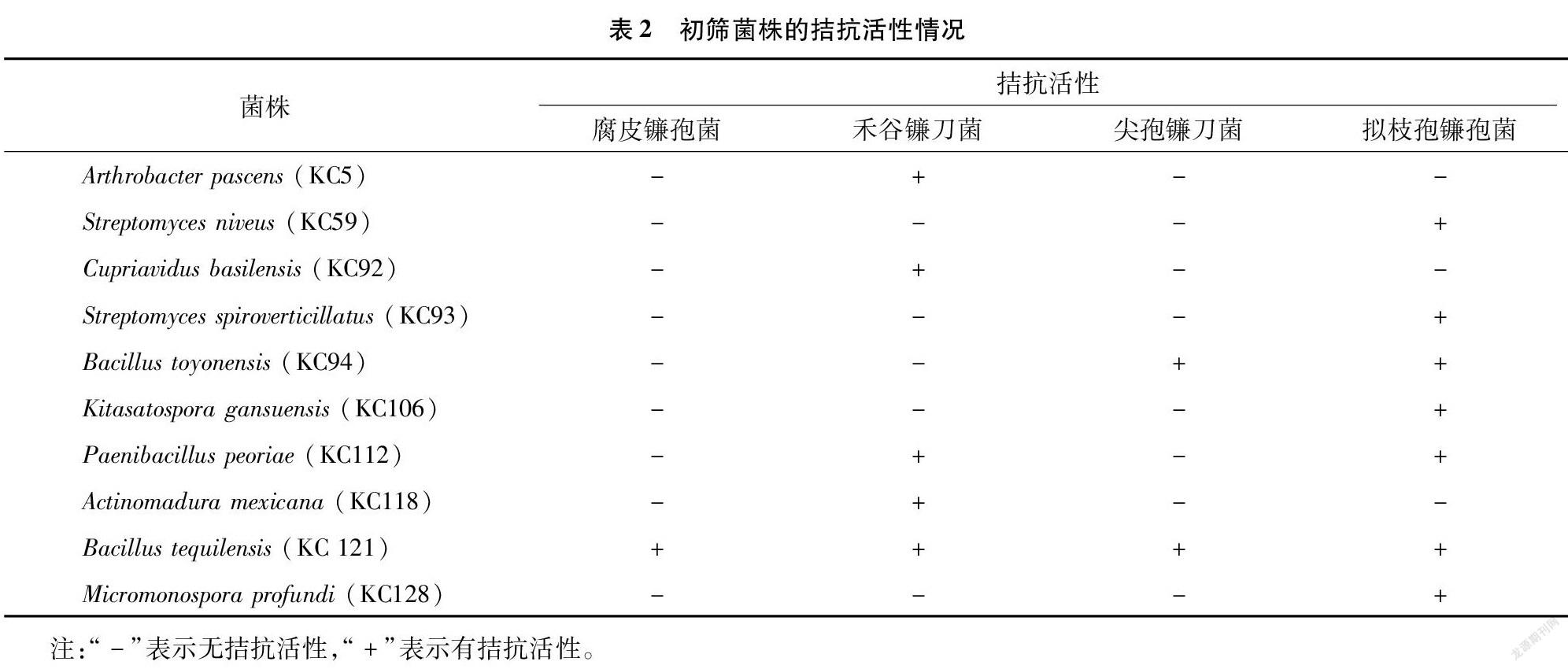
2.1 拮抗菌的初筛
从前期分离获得的83株菌株中,筛选得到10株具有不同程度抑制根腐病病原菌的菌株(表2)。其中,拮抗腐皮镰孢菌的生防菌最少,仅有1株;而拮抗拟枝孢镰孢菌和禾谷镰刀菌的生防菌相对较多,分别有7株和5株。KC 94和KC 112对2种病原菌的生长有拮抗作用,其余的7株供试菌株仅对1种病原菌有抑制效果。仅有KC 121对4种病原菌都具有拮抗活性,因此选择KC 121进行后续的复筛工作。
2.2 拮抗菌的复筛
2.2.1 拮抗菌KC 121对病原菌生长的影响
如图1所示,KC 121对根腐病的4种病原菌都有不同程度的抑制作用。其中,除对尖孢镰刀菌的抑制率较低外,对腐皮镰孢菌、禾谷镰刀菌和拟枝孢镰孢菌的抑菌率均高于50%,抑制效果最为明显的是拟枝孢镰孢菌,高达82.66%(表3)。
2.2.2 拮抗菌KC 121发酵液对病原菌生长的影响
病原菌在涂有发酵液的平板上培养7 d后观察发现(表4),KC 121发酵液对腐皮镰孢菌和拟枝孢镰孢菌的抑制作用较好,抑菌率分别为60.85%和8476%;而对禾谷镰刀菌和尖孢镰刀菌的抑制作用则相对较差,均不到50%,抑菌率最低的为9.20%。
2.3 拮抗菌KC 121的鉴定
2.3.1 形态特征 KC 121在ISP 2培养基中 28 ℃、培养16 h后观察发现,菌落呈浅肉色,边缘不规则且颜色浅于中央,表面褶皱、干燥(图2)。
2.3.2 生理生化特性 如表5所示,KC 121是一株具有接触酶和脲酶活性的菌株,能够消化的脂肪酸链长度为60,具有液化明胶、胨化牛奶、水解淀粉和还原硝酸盐的功能,不具备分解纤维素和产生硫化氢的能力。
2.3.3 分子生物学鉴定 菌株KC 121经16S rRNA基因测序后得到大小为1371bp的序列片段,将其提交到GenBank获得16S rDNA序列登录号为MW652665,同时在EzBioCloud (https://www.ezbiocloud.net/identify)进行比对, 用 MEGA 7软件采用邻接法(Neighbor-Joining)构建系统发育树[22]。KC 121的16S rRNA基因序列与Bacillus tequilensis(KF036187和MG004174)在系统发育树上同属一个分支,证明KC 121与特基拉芽孢杆菌(Bacillus tequilensis)的亲缘关系最近(图3)。
3 结论与讨论
利用笔者所在课题组前期从兰坪铅锌尾矿样品中分离得到的83株菌进行根腐病病原菌的生防菌筛选研究,初筛获得10株对不同病原菌具有不同抑制效果的生防菌,从中选择了1株能同时抑制腐皮镰刀菌、禾谷镰刀菌、尖孢镰刀菌和拟枝孢镰刀菌4种病原菌的菌株KC 121进行后续的复筛试验。本试验借鉴医学和药理学等研究中的平板涂布法[18,23],首次将菌株的发酵液涂布于平板上进行农业病害拮抗菌的筛选。通过平板对峙法和發酵液涂布法2种方法进行复筛,发现KC 121和其发酵液对拟枝孢镰孢菌的抑制效果均较好,抑菌率均达80%以上;但对尖孢镰刀菌的抑制效果相对较差,抑菌率均低于50%。在针对禾谷镰刀菌时,KC 121的菌体本身对病原菌菌丝生长的抑菌率为65.31%,但发酵液对其的抑菌率仅有9.20%,出现该情况的原因可能是由于抑菌物质为胞内产物,或是发酵条件(温度、培养基、时间和pH值等)不适宜所导致的。
结合形态学、生理生化试验检测及分子生物学测定,鉴定KC 121菌株为特基拉芽孢杆菌。多项研究报道表明,特基拉芽孢杆菌在诸多方面都有十分重要的作用与应用价值。在农业上,Cuellar等发现特基拉芽孢杆菌产生的脂肽能够显著降低香蕉叶斑病发生的严重性[24];周瑚等研究发现特基拉芽孢杆菌对多种植物病原菌有抑制作用,例如稻瘟病菌、辣椒胶孢炭疽菌、烟草赤星病菌、黄瓜疫病菌和卵菌具有一定的抑制作用[22]。在工业上,Angural等将特基拉芽孢杆菌应用在纸浆漂白中[25];Singh等研究发现特基拉芽孢杆菌能够产生生物表面活性剂,该活性剂可作为一种消毒剂或类似消毒剂的成分,对浮游细菌和生物膜中滞留的细菌均有杀菌作用[26]。在食品加工过程中,具有果胶酶活性的特基拉芽孢杆菌具备去除咖啡种子浆层的能力,部分纯化的果胶酶具有澄清果汁的作用[27]。而本研究首次发现了特基拉芽孢杆菌能够同时对镰刀菌属的腐皮镰刀菌、禾谷镰刀菌、尖孢镰刀菌和拟枝孢镰刀菌4种病原菌生长起拮抗作用,其中对拟枝孢镰孢菌的抑制效果最好,抑菌率达80%以上,为后期运用生物手段防治该病害提供了菌种资源。
菌株KC 121来源于重金属含量较高的尾矿极端环境。目前已有大量研究證明极端环境中存在丰富的防治农业病虫害的微生物资源。王彦等在河西走廊敦煌地区的盐碱土壤中分离筛选出1株链霉菌属菌株对黄瓜枯萎病病菌有良好的拮抗效果[28];闫建芳等在连黄海海域海底沉积物样品中获得1株密旋链霉菌(Streptomyces pactum)对番茄溃疡病菌(Clavibacter michiganensis)有较好的抑制作用[29];郑梅霞等从高海拔的可可西里和西藏样品中筛选得到2株能抑制番茄根腐病病原菌的枯草芽孢杆菌[30];罗曼等在南极沉积物样品中获得1株具有广谱抑菌作用的枯草芽孢杆菌斯氏亚种(Bacillus subtilis subsp. spizizenii),对丝瓜和辣椒的枯萎病病菌均有较强的抑制效果,对长豆褐腐病也有较明显的拮抗作用[31]。针对尾矿地区微生物的研究主要集中在多样性分析及重金属的污染生态修复等方面[32-36],尚未发现农业生防菌的相关研究和报道。本研究首次从含有大量重金属的兰坪铅锌尾矿极端环境中筛选生防菌,获得了活性较好的菌株,但抑菌物质、抑菌机理等方面还尚未明确,后续还需要进行更为深入的试验及研究。
参考文献:
[1]贺运春. 真菌学[M]. 北京:中国林业出版社,2008.
[2]张素轩. 镰刀菌属分类进展[J]. 真菌学报,1991,10(2):85-94.
[3]周 默,白庆荣. 内蒙古莫旗黄芪根腐病病原的分离与鉴定[J]. 东北农业科学,2021,46(2):52-55,61.
[4]张俊庆,郑安科,高立红,等. 辣椒枯萎病主要致病菌的分离鉴定及农用拮抗菌筛选[J]. 安徽农业科学,2021,49(8):134-137.
[5]赵东晓,陈 悦,夏 博,等. 沙棘枝干枯萎病症状观测与病原菌室内化学药剂筛选[J]. 中国水土保持,2021(4):50-53.
[6]王彤彤,邬昊月,赵尊练,等. 陕西省大葱叶部镰刀菌枯萎病病原菌的鉴定[J/OL]. 植物病理学报(2021-04-01)[2021-04-20].https://doi.org/10.13926/j.cnki.oppd.000568.
[7]De Oliveira R L,Reis G M,Da Silva V N,et al. Molecular characterization and fumonisin production by Fusarium verticillioides isolated from corn grains of different geographic origins in Brazil[J]. International Journal of Food Microbiology,2011,145(1):9-21.
[8]林镇跃,阙友雄,刘平武,等. 植物致病镰刀菌的研究进展[J]. 中国糖料,2014(1):58-64,78.
[9]刘利佳,李芳芳,何 雷,等. 烟草镰刀菌根腐病病原菌的鉴定及其对5种杀菌剂的敏感性分析[J]. 河南农业科学,2021,50(7):101-109.
[10]王国荣,冯晓晓,吴慧明,等. 芹菜茎基腐病病原菌鉴定、消长动态调查与防治药剂筛选[J]. 浙江农业学报,2021,33(4):661-669.
[11]肖荣凤,陈燕萍,陈梅春,等. 太子参根腐病病原菌的鉴定及防治药剂筛选[J]. 植物保护学报,2020,47(6):1333-1342.
[12]许 乐,王子强,张 爽,等. 丹参根腐病拮抗细菌筛选、鉴定及生防机理研究[J]. 中国生物防治学报,2021,37(4):846-854.
[13]牛世全,李 静,张雪莹,等. 一株抗黄芪根腐病芽孢杆菌的筛选、鉴定及抑菌物质的初步研究[J]. 西北师范大学学报(自然科学版),2021,57(2):79-86.
[14]程先锋,宋婷婷,陈 玉,等. 滇西兰坪铅锌矿区土壤重金属含量的高光谱反演分析[J]. 岩石矿物学杂志,2017,36(1):60-69.
[15]樊炳君,曹艳茹,纪开娟,等. 桉树根际放线菌的分离、初步鉴定及酶活筛选[J]. 昆明学院学报,2020,42(3):64-70.
[16]樊炳君,赵玉美,陈俊珠,等. 云南核桃叶枯病病原鉴定及其生物学特性[J]. 植物保护,2020,46(3):123-130.
[17]马 腾,张知晓,户连荣,等. 一株核桃真菌病害生防菌的鉴定及抑菌特性研究[J]. 中国森林病虫,2020,39(6):10-15.
[18]宋剑武,王鹏霞,吴永继,等. 迷迭香酸与抗菌药联合对含fosA3基因细菌抑菌效果的研究[J]. 中国畜牧兽医,2015,42(7):1851-1858.
[19]许丽婷,陈佳欣,李欢欢,等. 生防菌XC-1的筛选、鉴定及其对马铃薯黑痣病的防效研究[J]. 植物病理学报,2021,51(3):413-422.
[20]东秀珠,蔡妙英. 常见细菌系统鉴定手册[M]. 北京:科学出版社,2001:353-398.
[21]Marchesi J R,Sato T,Weightman A J,et al. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA[J]. Applied and Environmental Microbiology,1998,64(2):795-799.
[22]周 瑚,邹秋霞,胡 玲,等. 特基拉芽孢杆菌JN-369的分离鉴定及其抑菌物质分析[J]. 农药学学报,2019,21(1):52-58.
[23]刘 波,李雪驼,徐和利,等. 5种中药制剂杀灭幽门螺杆菌的实验研究[J]. 中国新药杂志,2002,11(6):457-459.
[24]Cuellar G T Z,González J L M,Villegas E V. Role of Bacillus tequilensis EA-CB0015 cells and lipopeptides in the biological control of black Sigatoka disease[J]. Biological Control,2021,155(4):104523.
[25]Angural S,Kumar A,Kumar D,et al. Lignolytic and hemicellulolytic enzyme cocktail production from Bacillus tequilensis LXM 55 and its application in pulp biobleaching[J]. Bioprocess and Biosystems Engineering,2020,43(12):2219-2229.
[26]Singh A K,Sharma P. Disinfectant-like activity of lipopeptide biosurfactant produced by Bacillus tequilensis strain SDS21[J]. Colloids and Surfaces B-Biointerfaces,2020,185:110514.
[27]Koshy M,De S. Effect of Bacillus tequilensis SALBT crude extract with pectinase activity on demucilation of coffee beans and juice clarification[J]. Journal of Basic Microbiology,2019,59(12):1185-1194.
[28]王 彥,牛世全,郑豆豆,等. 黄瓜枯萎病拮抗放线菌的筛选、鉴定及发酵条件优化[J]. 微生物学通报,2019,46(5):1062-1073.
[29]闫建芳,刘 秋,赵柏霞,等. 番茄溃疡病生防菌YH23的发酵条件优化及菌种鉴定[J]. 沈阳农业大学学报,2019,50(5):608-613.
[30]郑梅霞,朱育菁,刘 波,等. 高海拔来源芽胞杆菌根腐生防菌的筛选与鉴定[J]. 福建农业学报,2017,32(9):996-1000.
[31]罗 曼,万婧倞,黄仕新,等. 南极沉积物来源抗菌细菌的筛选及抑菌物质的鉴定[J]. 微生物学通报,2020,47(6):1787-1794.
[32]Njoku K L,Akinyede O R,Obidi O F. Microbial remediation of heavy metals contaminated media by Bacillus megaterium and Rhizopus stolonifer[J]. Scientific African,2020,10:1-17.
[33]杨雍康,药 栋,李 博,等. 微生物群落在修复重金属污染土壤过程中的作用[J]. 江苏农业学报,2020,36(5):1322-1331.
[34]彭玙萍,曾伟民. 紫金山铜矿酸性矿山废水微生物群落多样性[J]. 微生物学通报,2020,47(9):2887-2896.
[35]李司宇,刘 雪,王文婧,等. 微生物在重金属离子污染修复及治理中的应用研究[J]. 环境与可持续发展,2020,45(2):158-160.
[36]房保柱,王怡欢,张 堃,等. 凡口铅锌矿酸性底泥可培养微生物资源的探索分离[J]. 微生物学杂志,2020,40(2):10-21.
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2021年21期)2021-11-26
现代畜牧科技(2021年9期)2021-10-13
职工法律天地·下半月(2016年10期)2016-11-30
资治文摘(2016年7期)2016-11-23
农业与技术(2016年15期)2016-11-09
江苏农业科学(2016年3期)2016-05-03
上海蔬菜(2016年5期)2016-02-28
江苏农业科学(2015年5期)2015-10-20
天津农业科学(2014年9期)2015-05-06
