
Anle138b对慢性AβOs诱导海马神经元毒性作用影响
2021-11-17 02:23刘伟
青岛大学学报(医学版) 2021年5期
刘伟
[摘要] 目的 觀察二苯基吡唑化合物Anle138b对β淀粉样蛋白寡聚体(AβOs)诱导的海马神经元毒性作用的影响。
方法 原代培养的海马神经元细胞成熟后,将其分为空白对照组、AβOs处理组(用0.1 μmol/L AβOs处理7 d)和Anle138b处理组(用0.1 μmol/L AβOs处理7 d,在AβOs处理的后2 d加用0.1 μmol/L Anle138b)。使用乳酸脱氢酶(LDH)试剂盒检测细胞损伤情况,采用蛋白质免疫印迹(Western blot)法检测凋亡蛋白cleaved caspase-3的表达。
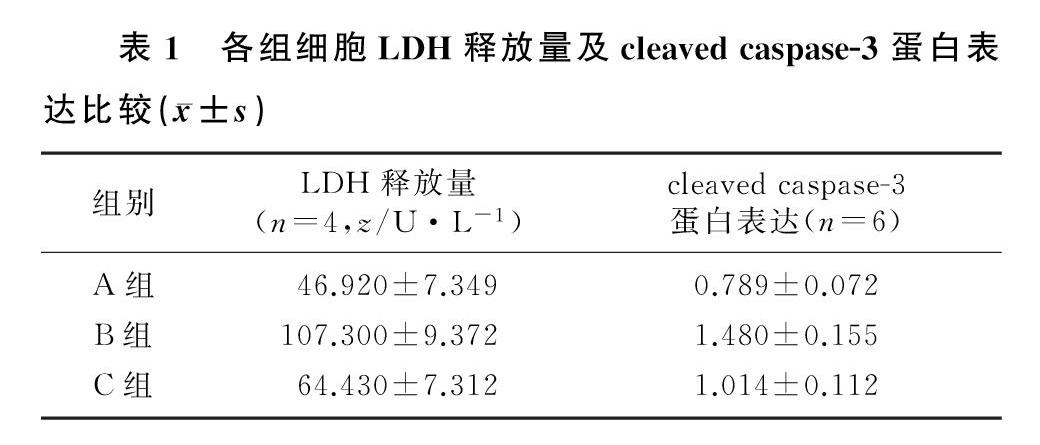
结果 与空白对照组相比较,AβOs处理组海马神经元细胞LDH释放量增加(F=14.810,q=7.481,P<0.01),凋亡蛋白cleaved caspase-3的表达升高(F=6.677,q=4.816,P<0.05)。与AβOs处理组相比,Anle138b处理组海马神经元细胞LDH释放量降低(q=5.310,P<0.05),凋亡蛋白cleaved caspase-3的表达有下降趋势,但差异无统计学意义(P>0.05)。
结论 Anle138b在慢性AβOs诱导的海马神经元凋亡模型中具有一定的保护作用。
[关键词] 阿尔茨海默病;蛋白质聚集体;淀粉样β肽类;Anle138b;海马;神经元;毒性作用
[中图分类号] R338.2
[文献标志码] A
[文章编号] 2096-5532(2021)05-0662-04
doi:10.11712/jms.2096-5532.2021.57.185
[开放科学(资源服务)标识码(OSID)]
[网络出版] https://kns.cnki.net/kcms/detail/37.1517.R.20211101.1333.003.html;2021-11-02 14:23:02
EFFECT OF ANLE138B ON CHRONIC HIPPOCAMPAL NEURONAL TOXICITY INDUCED BY β-AMYLOID PEPTIDE OLIGOMERS
LIU Wei
(Department of Physiology and Pathophysiology, School of Basic Medical, Qingdao University Medical College, Qingdao 266071, China)
[ABSTRACT] Objective To investigate the effect of the diphenyl-pyrazole compound Anle138b on hippocampal neuronal toxicity induced by β-amyloid peptide oligomers (AβOs).
Methods After the primary cultured hippocampal neurons became mature, they were divided into blank control group, AβOs treatment group (treated with 0.1 μmol/L AβOs for 7 days), and Anle138b treatment group (treated with 0.1 μmol/L AβOs 5 days and then treated with both AβOs and 0.1 μmol/L Anle138b for 2 days). Lactate dehydrogenase (LDH) kit was used to observe cell damage, and Western blot was used to measure the expression of the apoptosis protein cleaved caspase-3.
Results Compared with the blank control group, the AβOs treatment group had significant increases in the release of LDH in hippocampal neurons (F=14.810,q=7.481,P<0.01) and the expression of the apoptotic protein cleaved caspase-3 (F=6.677,q=4.816,P<0.05). Compared with the AβOs treatment group, the Anle138b treatment group had a significant reduction in the release of LDH in hippocampal neurons (q=5.310,P<0.05), as well as a tendency of reduction in the expression of the apoptotic protein cleaved caspase-3, but with no significant difference (P>0.05).
Conclusion
Anle138b has a certain protective effect in the model of chronic hippocampal neuronal apoptosis induced by AβOs.
[KEY WORDS] Alzheimer disease; protein aggregates; amyloid beta-peptides; Anle138b; hippocampus; neurons; toxic actions
阿尔茨海默病(AD)又称为老年痴呆症,是一种多发于老年人群的神经退行性疾病,也是痴呆症最常见的病因。随着预期寿命的持续增长,AD发病率将会越来越高。到2050年,全世界痴呆症的人数预计将增加到1.315亿[1]。尽管进行了数十年的研究,但该病至今尚未找到有效的治疗方法。AD的病因复杂,涉及遗传、环境等多种因素。目前关于AD病因的假说有很多,包括胆碱能假说、β淀粉样蛋白(Aβ)毒性假说、tau蛋白异常修饰假说、钙稳态假说等,其中被广泛认同的是Aβ毒性假说[2-3]。离子通道假说是Aβ发挥毒性的一种机制,该假说认为,Aβ变构、组装成离子通道結构,嵌入脂质双层膜,造成膜泄漏和钙稳态失衡,进而导致神经元的损伤和死亡[4-5]。
Anle138b是一种低聚物调节剂,具有出色的口服生物利用度和血-脑脊液屏障渗透性,并且在治疗剂量下没有可检测到的毒性[6]。分子动力学模拟实验表明,Anle138b可以有效地阻断肽链相互作用并阻止自发形成有序的β-折叠结构[7]。2018年,MAR-TINEZ HERNANDEZ等[8]对AD大鼠模型及人工膜的研究表明,Anle138b可以阻止Aβ打孔的活动,而不会改变膜嵌入的Aβ低聚物结构。Anle138b是首个被报道的能够在通道形成后将其阻断并改善Aβ病理变化的化合物,并且还可以减少病理性tau蛋白聚集体,是治疗AD相关病理改变的新型且有希望的化合物,但是该药在细胞模型上的研究结果较少[9]。因此,本实验在细胞模型上通过对海马神经元凋亡损伤情况的检测,观察了Aβ通道抑制剂Anle138b对慢性Aβ寡聚体(AβOs)诱导的海马神经元毒性作用的影响,为开发治疗AD新型药物提供思路。现将结果报告如下。
1 材料与方法
1.1 主要材料
原代培养的海马神经元细胞(从大鼠新生鼠脑内获取),DMEM/F12基础培养液、胰蛋白酶(美国Hyclone公司),胎牛血清(FBS,美国Gibco公司),Anle138b(Medchem Express公司),青霉素-链霉素溶液(100×,北京索莱宝科技有限公司),Aβ42单体粉末(杭州丹港生物科技有限公司),BCA蛋白定量检测试剂盒(武汉赛维尔生物科技有限公司),D-多聚赖氨酸(美国Sigma公司),cleaved caspase-3抗体(上海Cell Signaling公司),ECL发光液以及PAGE凝胶快速制备试剂盒(上海雅酶生物科技有限公司),乳酸脱氢酶(LDH)检测试剂盒(南京建成生物工程研究所)。
1.2 实验方法
1.2.1 AβOs制备 取1 mg的Aβ42粉末和六氟异丙醇(HFIP)置于冰上预冷,向装有Aβ42粉末的EP管中加入222 μL的HFIP,密封,涡旋混匀,室温下孵育60 min,直到液体澄清,得到1 mmol/L的 Aβ-HFIP溶液。将Aβ-HFIP溶液置于冰上5 min,取4只无菌EP管,分装Aβ-HFIP溶液55 μL,在通风橱中将HFIP挥干,得到无色透明Aβ肽膜,置-20 ℃冰箱保存。使用前,取1支分装管,在冰上加入二甲基亚砜(DMSO)11 μL,水浴超声(300 W,35 Hz)处理10 min,加入磷酸盐缓冲液(PBS)539 μL,涡旋混匀,置于冰箱中4 ℃孵育1 d。使用离心机在4 ℃下以13 000 r/min离心10 min,上清即为100 μmol/L的AβOs[10]。
1.2.2 海马神经元细胞培养 实验前先将所有实验器械高压灭菌以确保细胞不被污染。12孔板用多聚赖氨酸包被,在细胞培养箱中放置2 h以上,实验前用高压后的双蒸水冲洗3次,去除多聚赖氨酸,放于培养箱中备用。将小鼠浸泡于体积分数0.75的乙醇中消毒,用剪刀剪下头部后取出大脑,将其置于盛有基础培养液预冷的培养皿中,在解剖显微镜下取出海马体(位于大脑皮质内,有较为明显的血管膜界限),并去除血管膜。加入胰蛋白酶5 mL,在细胞培养箱中消化5 min,使用含有FBS的完全培养液终止消化。使用移液枪将海马体移至新的培养皿中,将其吹打均匀,沉淀后转移上清至离心管中,以1 000 r/min离心5 min,弃上清,用含B27的培养液轻轻吹打混匀。用细胞计数板进行细胞计数,调整细胞密度约为7×108/L。将细胞悬液以每孔1.5 mL接种于6孔板中,置于37 ℃、含体积分数0.05 CO2的培养箱中培养,每隔3 d进行1次半换液,细胞培养7~8 d后观察细胞状态,待神经元胞体发育饱满、突触连接明显后用于后续实验。
1.2.3 实验分组及处理 实验分为空白对照组(A组,无处理措施)、AβOs处理组(B组,用0.1 μmol/L的AβOs处理7 d)和Anle138b处理组(C组,用0.1 μmol/L的AβOs处理7 d,在AβOs处理后5 d加用0.1 μmol/L的Anle138b)。
1.2.4 海马神经元LDH释放量检测 细胞处理结束后,转移各组上清至EP管中,并置于冰上。按照试剂盒说明书进行LDH检测,操作过程应谨慎迅速,防止LDH降解。将待测样本转移到96孔板中,加入基质缓冲液和辅酶Ⅰ后,37 ℃孵育15 min;加入2,4-二硝基苯肼,37 ℃孵育15 min;加入NaOH溶液,室温孵育5 min;用酶标仪测定各孔在450 nm波长处的吸光度值,按照试剂盒说明书的公式计算培养液中LDH含量。
1.2.5 蛋白质免疫印迹(Western blot)方法检测cleaved caspase-3的表达 将待测样本转移到12孔板中,每孔加入蛋白裂解液60 μL,在冰上裂解30 min,使用刮板将贴附在板底的海马神经元细胞刮净,转移裂解液和细胞至EP管中,在4 ℃下以12 000 r/min离心20 min,吸取上清至新的EP管中,使用BCA试剂盒测定样品蛋白浓度。加入1/4上清量的5×Loading Buffer,95 ℃加热5 min,样品置于-20 ℃保存。使用125 g/L的PAGE凝胶进行电泳,将制备好的蛋白样品加入胶孔中,以80 V电压跑浓缩胶,120 V电压跑分离胶。电泳结束后,取出凝胶,应用0.22 μm的PVDF膜进行转膜,以300 mA恒定电流在冰上转膜约90 min。转膜结束后,用封闭液封闭 2 h,根据分子量大小裁膜,得到相关蛋白条带后加入一抗(稀释比例1∶800),4 ℃摇床孵育过夜,TBST 洗3次,每次10 min;加入二抗(稀释比例1∶10 000)后室温孵育 1 h,TBST洗3次,每次10 min。ECL发光试剂显影后,用Image J软件进行灰度分析,结果以目的蛋白与内参照蛋白β-actin的灰度值之比表示。
1.3 统计学处理
采用GraphPad Prism 7软件进行统计学分析,实验结果以±s的形式表示,多组数据比较采用单因素方差分析,并继以Tukey法进行多重比较。以P<0.05为差异有统计学意义。
2 结 果
2.1 Anle138b对AβOs诱导细胞凋亡模型LDH释放量的影响
与空白对照组相比,AβOs处理组海马神经元细胞LDH释放量增加(F=14.810,q=7.481,P<0.01);与AβOs处理组相比较,Anle138b处理组海马神经元细胞的LDH释放量降低(q=5.310,P<0.05)。见表1。
2.2 Anle138b对AβOs诱导细胞凋亡模型cleaved caspase-3蛋白表達的影响
Western blot检测结果显示,与空白对照组相比,AβOs 处理组细胞凋亡蛋白cleaved caspase-3的表达升高(F=6.677,q=4.816,P<0.05);与AβOs处理组相比较,Anle138b处理组海马神经元细胞cleaved caspase-3蛋白的表达有下降趋势,但差异无统计学意义(q=3.255,P>0.05)。见表1。
3 讨 论
随着社会老龄化的加剧,在中老年人群中AD患病率成倍增加,给社会和家庭带来沉重负担。目前美国食品药品监督管理局批准的治疗AD药物主要有两类,一类是胆碱酯酶抑制剂,另一类是兴奋性氨基酸受体拮抗剂[11]。尽管已证明这些药物对AD症状的控制具有统计学意义,但它们的治疗效果并不稳定,并且作用持续时间有限[12]。基于现有疗法的低效率,迫切需要开发能阻止疾病发展的药剂,找到AD的新疗法。而Aβ通道小分子阻断剂将会是治疗AD相关病理改变的一种新颖且有希望的化合物[9]。Anle138b是首个被报道的能够在通道形成后将其阻断并改善Aβ病理变化的化合物,但是该药物在细胞模型上的研究结果较少,因此本实验使用体外培养的原代海马神经元作为实验材料,并且降低AβOs给药浓度,采用连续7 d慢性给药的方式,研究Anle138b对慢性AβOs诱导的海马神经元毒性作用的影响。相较于人工膜以及急性分离模型,原代培养的海马神经元可以形成较为发达的神经网络,细胞间可以进行突触传递和信号交流,更加接近生理环境。本实验先给予AβOs处理5 d,再用Anle138b和AβOs共处理2 d,这种给药方式更能表现出Anle138b能够在Aβ通道形成后将其阻断的作用机制,这也是本实验设计的创新性所在。
本文的实验结果显示,与AβOs处理组相比,Anle138b处理能够降低海马神经元细胞LDH释放量,并且凋亡蛋白cleaved caspase-3的表达也呈下降趋势。表明Anle138b在慢性AβOs诱导的海马神经元凋亡模型中有一定的保护作用。后续研究尚需进一步加大样本量,这样cleaved caspase-3蛋白表达量的下降将可能有统计学意义,实验结果会更明确且具有说服力。近年来,Aβ如何影响神经元和突触功能受到关注[13-15]。而Aβ与质膜的相互作用是AD发展的关键,二者相互作用的结果之一就是形成离子通道[16-18]。离子通道形成后,会造成膜泄漏和钙稳态失衡进而导致神经元的损伤和死亡[19]。本研究Anle138b表现出对海马神经元的保护作用可能就是通过阻断Aβ通道的作用。到目前为止,在包括轻度至中度AD病人的临床试验中,大部分治疗方法均未成功通过Ⅲ期临床试验,进行分析时,在疾病过程中采取干预措施开始太晚是解释此类失败的经常性的论据之一,认为早期的预防性治疗将带来好处[20]。aducanumab抗体的临床试验也是集中在早期或中期病人[21-23],然而实际上AD病人被确诊基本处于发病后期,并且目前AD发病早期的诊断尚不成熟[24]。因此,寻求一种能在AD发病后期发挥重要作用的药物非常重要。而在本实验中,Anle138b加药时间是在AβOs处理一定时间之后,故像Anle138b这种小分子阻断剂可能在该病的治疗中有很大的潜力[25]。
[参考文献]
[1]SENGOKU R. Aging and Alzheimers disease pathology[J]. Neuropathology, 2020,40(1):22-29.
[2]LIU P P, XIE Y, MENG X Y, et al. History and progress of hypotheses and clinical trials for Alzheimers disease[J]. Signal Transduction and Targeted Therapy, 2019,4:29.
[3]LANE C A, HARDY J, SCHOTT J M. Alzheimers disease[J]. European Journal of Neurology, 2018,25(1):59-70.
[4]SHIRWANY N A, PAYETTE D, XIE J, et al. The amyloid beta ion channel hypothesis of Alzheimers disease[J]. Neuropsychiatric Disease and Treatment, 2007,3(5):597-612.
[5]ARISPE N, POLLARD H B, ROJAS E. Giant multilevel
cation channels formed by Alzheimer disease amyloid beta-
protein[A beta P-(1-40)] in bilayer membranes[J]. PNAS, 1993,90(22):10573-10577.
[6]WAGNER J, RYAZANOV S, LEONOV A, et al. Anle138b: a novel oligomer modulator for disease-modifying therapy of neurodegenerative diseases such as prion and Parkinsons di-
sease[J]. Acta Neuropathologica, 2013,125(6):795-813.
[7]LEVIN J, SCHMIDT F, BOEHM C, et al. The oligomer modulator anle138b inhibits disease progression in a Parkinson mouse model even with treatment started after disease onset[J]. Acta Neuropathologica, 2014,127(5):779-780.
[8]MARTINEZ HERNANDEZ A, URBANKE H, GILLMAN A L, et al. The diphenylpyrazole compound anle138b blocks Aβ channels and rescues disease phenotypes in a mouse model for amyloid pathology[J]. EMBO Molecular Medicine, 2018,10(1):32-47.
[9]GUIX F X, DOTTI C G. Could blocking the formation of amyloid channels rescue Alzheimers phenotype[J]? EMBO Molecular Medicine, 2018,10(1):7-9.
[10]CIUDAD S, PUIG E, BOTZANOWSKI T, et al. Aβ(1-42) tetramer and octamer structures reveal edge conductivity pores as a mechanism for membrane damage[J]. Nature Communications, 2020,11(1):3014.
[11]BRIGGS R, KENNELLY S P, ONEILL D. Drug treatments in Alzheimers disease[J]. Clinical Medicine (London, England), 2016,16(3):247-253.
[12]PANZA F, LOZUPONE M, DIBELLO V, et al. Are antibo-
dies directed against amyloid-β (Aβ) oligomers the last call for the Aβ hypothesis of Alzheimers disease[J]? Immunotherapy, 2019,11(1):3-6.
[13]FABIANI C, ANTOLLINI S S. Alzheimers disease as a membrane disorder: spatial cross-talk among beta-amyloid peptides, nicotinic acetylcholine receptors and lipid rafts[J]. Frontiers in Cellular Neuroscience, 2019,13:309.
[14]ARBOR S C, LAFONTAINE M, CUMBAY M. Amyloid-beta Alzheimer targets-protein processing, lipid rafts, and amyloid-beta pores[J]. The Yale Journal of Biology and Medicine, 2016,89(1):5-21.
[15]RANGACHARI V, DEAN D N, RANA P, et al. Cause and consequence of Aβ-Lipid interactions in Alzheimer disease pathogenesis[J]. Biochimica et Biophysica Acta Biomembranes, 2018,1860(9):1652-1662.
[16]POPUGAEVA E, PCHITSKAYA E, BEZPROZVANNY I. Dysregulation of intracellular calcium signaling in Alzheimers disease[J]. Antioxidants & Redox Signaling, 2018,29(12):1176-1188.
[17]VETRIVEL K S, THINAKARAN G. Membrane rafts in Alzheimers disease beta-amyloid production[J]. Biochimica et Biophysica Acta, 2010,1801(8):860-867.
[18]DI SCALA C, CHAHINIAN H, YAHI N, et al. Interaction of Alzheimers β-amyloid peptides with cholesterol: mechanistic insights into amyloid pore formation[J]. Biochemistry, 2014,53(28):4489-4502.
[19]BEZPROZVANNY I, MATTSON M P. Neuronal calcium mishandling and the pathogenesis of Alzheimers disease[J]. Trends in Neurosciences, 2008,31(9):454-463.
[20]ROSENTHAL J, BELFORT G, ISAACSON D. Early treatment critical: bexarotene reduces amyloid-beta burden in silico[J]. PLoS One, 2016,11(4):e0153150.
[21]ARNDT J W, QIAN F, SMITH B A, et al. Structural and kinetic basis for the selectivity of aducanumab for aggregated forms of amyloid-β[J]. Scientific Reports, 2018,8(1):6412.
[22]GAMAGE K K, KUMAR S. Aducanumab therapy ameliorates calcium overload in a mouse model of Alzheimers di-
sease[J]. The Journal of Neuroscience, 2017,37(17):4430-4432.
[23]KASTANENKA K V, BUSSIERE T, SHAKERDGE N, et al. Immunotherapy with aducanumab restores calcium homeostasis in Tg2576 mice[J]. The Journal of Neuroscience, 2016,36(50):12549-12558.
[24]BATEMAN R J, XIONG C J, BENZINGER T L, et al. Clinical and biomarker changes in dominantly inherited Alzhei-
mers disease[J]. The New England Journal of Medicine, 2012,367(9):795-804.
[25]MATTHES D, GAPSYS V, GRIESINGER C, et al. Resolving the atomistic modes of Anle138b inhibitory action on peptide oligomer formation[J]. ACS Chemical Neuroscience, 2017,8(12):2791-2808.
(本文編辑 马伟平)
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
中国药学药品知识仓库(2021年11期)2021-11-25
保健与生活(2020年6期)2020-03-20
中小学德育(2020年11期)2020-03-18
恋爱婚姻家庭·养生版(2019年1期)2019-01-25
中国卫生产业(2018年12期)2018-05-14
教育界·上旬(2016年12期)2017-05-25
环球人文地理(2016年10期)2016-11-24
科技知识动漫(2016年6期)2016-06-24
汽车观察(2009年1期)2009-02-18
