
典型温带针阔叶混交林碳收支对温度的响应特征
2021-11-15 07:37舒子倩刘勋埙李舒颖
中低纬山地气象 2021年5期
舒子倩,刘勋埙,姚 宁,吴 虑,李舒颖
(贵州省剑河县气象局, 贵州 剑河 556400)
0 引言
自十八世纪以来,在工业活动的迅速发展下使得大量的化石燃料被开采和燃烧,这些人类活动导致大气中CO2等温室气体浓度不断上升,从而导致全球变暖,这将严重的影响到森林生态系统的碳循环过程[1-2]。森林作为陆地上面积最大的生态系统,温度变化会影响其光合作用、呼吸作用过程,使其与大气之间的碳交换发生变化,进而又影响到大气CO2浓度[3]。因此,在气候变暖的条件下,分析森林碳循环过程对温度的响应特征成为了焦点[4]。
森林生态系统的碳交换包括:生态系统总初级生产力(Gross primary productivity,GPP:单位地表面积上单位时间内绿色植物通过光合作用途径所固定的碳量)、生态系统呼吸(Ecosystem respiration,Re:单位地表面积单位时间生态系统所有有机体的呼吸总量,包括自养呼吸和异养呼吸)以及净生态系统碳交换(Net ecosystem exchange of CO2,NEE:大气—植被界面的净CO2交换量)[5]。有许多研究者对森林生态系统GPP、Re、NEE就温度的响应特征进行探索[2]。研究表明,在小时尺度上,长白山温带针阔混交林、千烟洲亚热带人工针叶林和鼎湖山亚热带常绿阔叶林森林生态系统的GPP、Re对5 cm土壤温度以及气温的响应均呈指数形式。对于生态系统净碳交换量NEE,在日尺度上,长白山温带针阔混交林NEE在日平均气温Ta<0 ℃时,生态系统有微小的CO2排放,Ta>0 ℃时,随着日平均气温上升该生态系统的净碳吸收量增加[6]。这些研究都明确的表明在不同时间尺度上生态系统碳交换对温度的响应特征存在差异,因此需完善不同时间尺度上长白山温带针阔叶混交林森林生态系统碳收支各组分对温度的响应特征。
长白山温带针阔叶混交林是我国温带地区典型的森林生态系统类型,位于东北样带的东端,对气候变化有着较高的敏感性[7,8]。本文以长白山温带针阔叶混交林为研究对象,在获取连续监测数据的基础上,通过分析,明确该生态系统总初级生产力GPP、生态系统呼吸Re以及净生态系统碳交换NEE在不同的时间尺度上对气温(Ta),5 cm土壤温度(Ts)的响应特征,从而了解该类森林生态系统碳循环对气候变化的响应及适应特征。
1 研究方法
1.1 研究地概况
长白山温带针阔混交林位于吉林省东南部(41°24′09″N、128°05′45″E)是我国面积最大、自然环境和生态系统保存最为完整的森林生态系统保护区之一[7]。研究区域属于温带大陆性季风气候,是保存尚好的亚洲东部典型的山地森林生态系统,生物区系复杂,生物多样且丰富。观测地点的主要乔木有红松、椴树、蒙古栎、水曲柳、色木等。下木覆盖率为40%,土壤为山地暗棕色森林土[2,8]。
1.2 数据来源
中国科学院长白山森林生态系统定位站通量观测塔上观测的2003—2009年常规气象数据(包括太阳辐射、降水、气温和土壤5 cm处温度)以及经过通量数据处理程序处理之后得到的NEE、GPP、Re数据。
1.3 NEE的观测方法
涡度相关法:根据涡度相关理论,CO2通量的表达式为[4]:

(1)
式中,ρ为空气密度,w为垂直风速,c为CO2浓度,上划线表示某时间段内的平均值,撇号表示瞬时值与平均值的偏差即脉动值。涡度相关观测系统包括三维超声风温仪(CSAT3,Campbell Scientific Inc.,Logan,UT,USA)和CO2/H2O红外气体分析仪(LI-7500,LICOR Inc.,Lincoln,NE,USA),观测的原始采样频率为10 Hz,通过数据采集器计算其30 min平均值并记录[2,6-8]。当在森林这种具有高大冠层的生态系统进行观测时,还要考虑涡度相关系统探头高度以下CO2浓度变化导致的CO2储存,因此净生态系统CO2交换量(NEE)为[2]:
NEE=FC+FS
(2)
式中,FC为测量的CO2通量,FS为CO2储存项。探头高度以下的大气CO2储存项FS利用CO2/ H2O 分析仪测定的CO2浓度随时间的变化进行计算[1,9,10]。NEE为矢量,其值为正代表CO2交换方向向上,生态系统释放CO2,其值为负代表CO2交换方向向下,生态系统吸收CO2[2]。
1.4 Re的观测及计算方法
涡度相关法观测 :涡度相关法直接观测的是NEE,但光合作用只发生在生长季白天,因此夜间观测的NEE即为生态系统呼吸,白天的生态系统呼吸可通过夜间呼吸外推法进行计算,即将夜间测定的有效NEE(即Re)通过温度响应模型外推到白天,从而求解白天的Re[2]。
在不受水分胁迫的影响下,生态系统呼吸对温度的响应特征可用Lloyd & Taylor方程进行描述:
(3)
式中Reco,ref是参考温度(Tref= 283.16 K)下的生态系统呼吸(单位:mg·m-2·s-1);TK为温度(单位:K);E0在实际应用中为 309 K;T0为温度试验常数(227.14 K)[2]。
1.5 GPP的估算方法
生态系统水平上,无法直接观测到GPP,主要通过计算生态系统呼吸Re以及净生态系统碳交换NEE来得到,即根据公式(4),通过观测的NEE以及计算的Re来得到[2]。
NEE=-GPP+Re
(4)
2 不同时间尺度上生态系统碳收支对温度的响应特征
2.1 不同时间尺度上GPP对温度的响应特征
由于2003—2008年的每年日尺度上GPP分别对日平均气温和日平均5 cm土壤温度的响应形式均为指数形式,我们在这里仅使用2003年的关系图来说明,其余年份的拟合指数方程的回归系数和R2如表1所示。而在月尺度上,我们通过长白山温带针阔叶混交林计算得到6 a每年12个月的月总GPP以及月平均气温Ta和月平均5 cm土壤温度Ts,做出2003—2008年该生态系统月尺度上GPP分别对月平均气温Ta和月平均5 cm土壤温度Ts的响应图。

表1 2003—2008年日尺度上GPP对日平均气温Ta、日平均5 cm处土壤温度Ts的响应趋势方程(y=aebx)系数与R2
根据图1我们可以看出,在日尺度和月尺度上长白山温带针阔叶混交林生态系统GPP对气温和5 cm土壤温度的响应呈指数形式。其趋势均为:当气温Ta<0 ℃时,GPP的变化不大;当Ta>0 ℃时,可以明显看出GPP随温度的增加而增大,碳吸收能力增强,但离散程度也增加。这是因为除了气温的影响外,GPP还与生态系统物候、光合有效辐射(PAR)、空气湿度等因素有关[2,9]。
与日尺度上GPP对气温和土壤温度的响应对比,我们可以看出在月尺度上的离散度比日尺度上的小,同时相关性也比日尺度上的大,这也说明了在月尺度上气温和土壤温度对GPP的影响作用更强,相关性更大。同时,在日尺度和月尺度上较GPP对气温的响应,长白山温带针阔叶混交林GPP对5 cm土壤温度的响应(图1b、1d)的离散程度较大。这是由于在5 cm土壤温度低于0 ℃时,由于温度太低,植被停止生长,GPP基本没有太大的变化,略大于0;当5 cm土壤温度高于0 ℃时,GPP就随其的升高而指数增大。
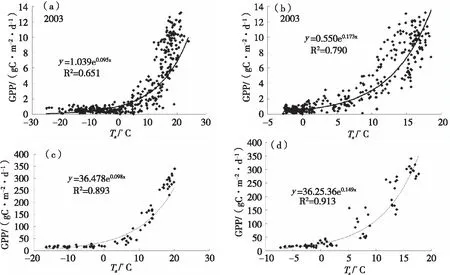
图1 长白山温带针阔叶混交林日尺度和月尺度上GPP随日(a)、月(c)平均气温Ta和日(b)、月(d)平均5cm土壤温度Ts的变化
根据以上在不同时间尺度上长白山温带针阔混交林生态系统GPP对温度(气温和5 cm土壤温度)的响应趋势的分析,我们发现日尺度和月尺度上该森林生态系统GPP对气温和土壤温度的响应均呈指数形式,这与其他学者在研究其他温带、亚热带森林生态系统在日尺度和月尺度上GPP对温度的响应的结果是一样的[1,10-12]。同时我们还发现,对于GPP与气温和5 cm土壤温度的相关性都很高,但趋势变化的离散程度也较大,这主要是由于受到太阳辐射、空气湿度、土壤含水量等因素的影响[2,6]。而温度低于0 ℃以后,生态系统的GPP仍为正值,这是由于长白山生态系统处于非生长季,植被处于休眠状态,光合作用十分微弱,但土壤中有微弱的微生物呼吸和分解作用释放的CO2。
2.2 不同时间尺度上Re对温度的响应特征
长白山温带针阔叶混交林生态系统的生态系统呼吸Re在日尺度和月尺度上与日气温和5 cm土壤温度(如图2)都呈指数关系,随温度(气温和土壤温度)的升高生态系统呼吸随之增加。当温度低于0 ℃时,生态系统呼吸强度很微弱,且随温度的升高,变化不大。当温度高于0 ℃时,随着温度的升高生态系统呼吸Re也逐渐增强。但同时离散程度也增大,这与日尺度上GPP对温度的响应十分相似,可能是由于生态系统的物候、空气湿度等因素的影响[6]。
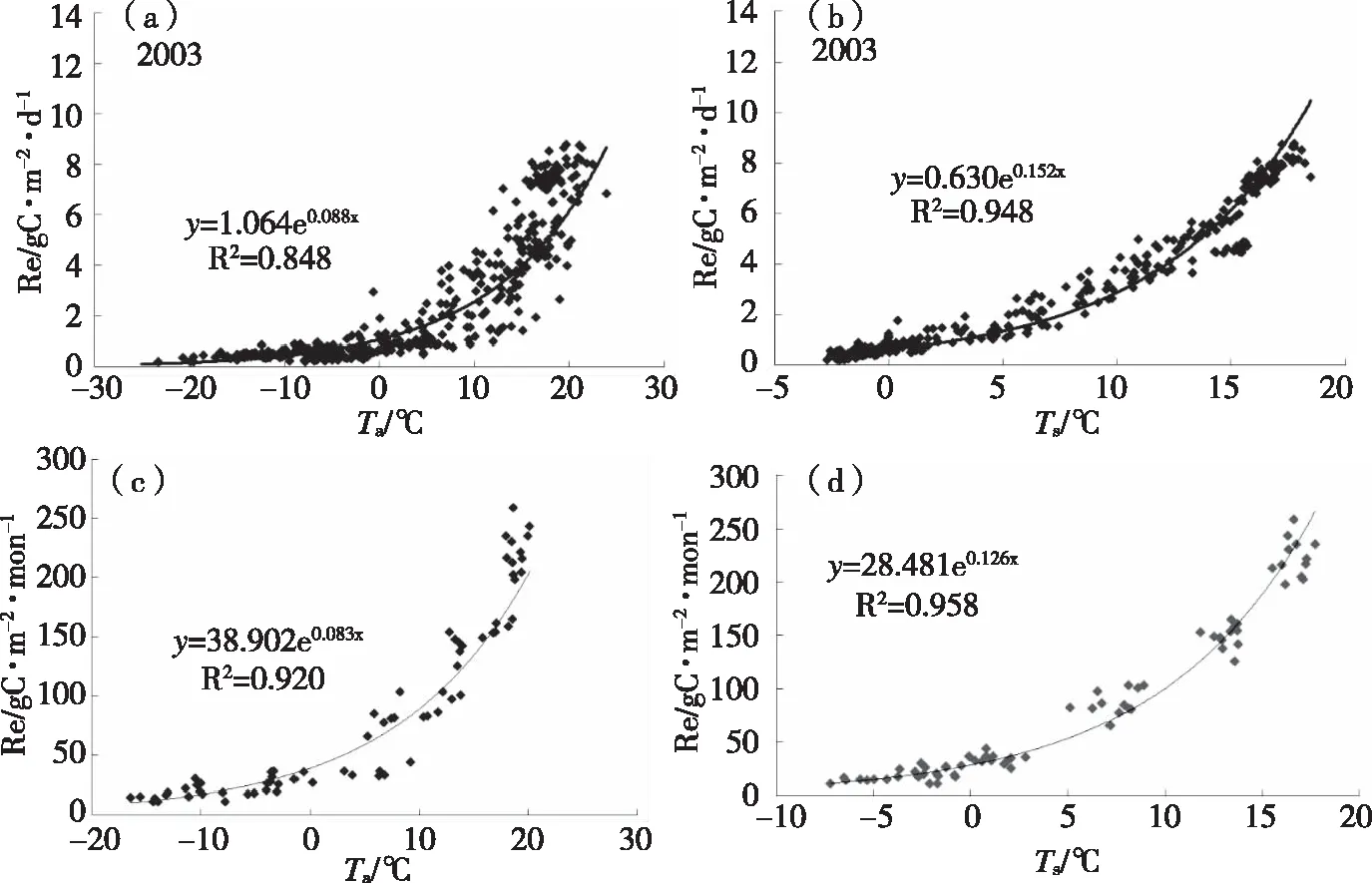
图2 长白山温带针阔叶混交林日尺度和月尺度上Re随日(a)、月(c)平均气温Ta和日(b)、月(d)平均5cm土壤温度Ts的变化
相比于对气温的响应,在日尺度和月尺度上Re与5 cm处土壤温度的相关程度更好,当土壤温度低于0 ℃时,Re的变化不大;当土壤温度高于0 ℃时,Re随土壤温度的增加而迅速增加,碳排放能力增强,同时,与Re对气温的响应的离散度相比更小,这也明显地说明了土壤温度是影响该生态系统呼吸的主要因素之一。并且对于长白山温带针阔混交林森林生态系统来说,其生态系统呼吸Re对5 cm土壤温度比对气温的相关性更好。
根据表2的2003—2008年日尺度上Re分别对日平均气温Ta和日平均5cm处土壤温度Ts的响应趋势方程(y=aebx)系数与R2可以发现,这6 a在日尺度上Re对日平均气温(Ta)和日平均5cm处土壤温度(Ts)的响应趋势均为显著的指数形式。
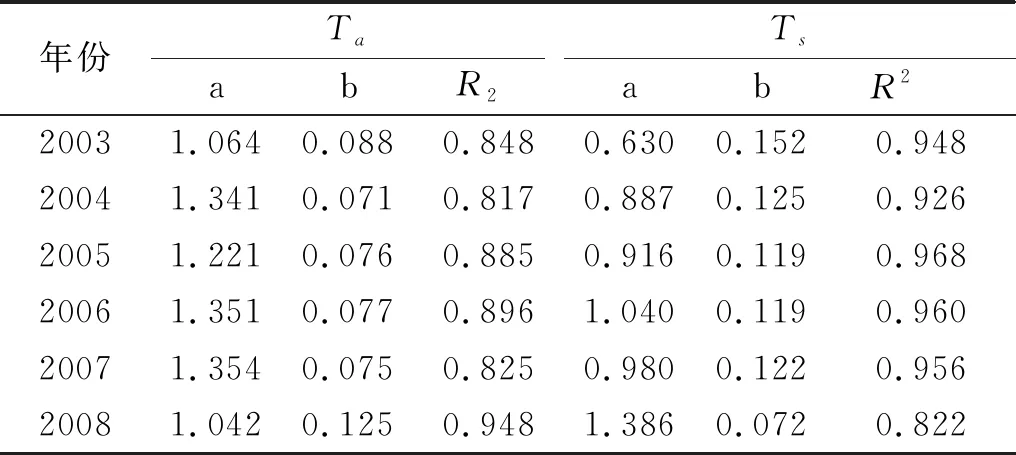
表2 2003—2008年日尺度上Re对日平均气温Ta、对日平均5cm处土壤温度Ts的响应趋势方程(y=aebx)系数与R2
通过对不同时间尺度的长白山温带针阔叶混交林生态系统Re对温度(气温和5 cm土壤温度)的响应分析可以得到,在日尺度和月尺度上,该生态系统Re随气温和5 cm土壤温度的变化均呈指数形式,且相关性均较好,但对于长白山温带针阔混交林森林生态系统来说,该生态系统的Re与5 cm土壤温度的相关性更好。这与其他学者的研究相同,与亚热带森林生态系统的呼吸与气温相关性更高相比,长白山温带针阔混交林森林生态系统是与5 cm土壤温度的相关性更好[6]。
2.3 不同时间尺度上NEE对温度的响应特征
从图3中可以看出,在日尺度和月尺度上,NEE对气温的响应均分为2个阶段,该生态系统NEE随气温的变化首先是随着气温的升高而逐渐向正的方向增大,即NEE随着日平均气温的升高,生态系统主要表现为碳排放。第2个阶段,当气温达到一定值时,日总NEE随气温的迅速增长而转变为向负的方向增大,即生态系统由碳排放转变为碳吸收。这主要是由于,当气温较低时生态系统基本没有光合作用,但是生态系统的呼吸作用依然在进行并且保持一定的强度,这导致了生态系统总体上表现为碳释放,并且随着气温的逐渐升高,生态系统呼吸作用所释放的CO2越来越多。而当气温达到一定的温度值时,生态系统光合作用吸收CO2的能力逐渐增强且大于呼吸作用所排放的碳,生态系统表现为碳吸收,并且碳吸收的强度随着温度的进一步升高而增强。同时,离散度也相应增大,这可能是由于太阳辐射,空气湿度等其他因素引起的。而在这里,达到一定温度水平后发生转折的这个温度,我们称之为碳源向碳汇转变的临界温度(T0)。

图3 长白山温带针阔叶混交林日尺度(a)和月尺度(b)上NEE分别随平均气温Ta的变化
同时还可以看出,在月尺度上NEE随气温的变化趋势中离散度较大,这说明在月尺度上由于GPP和Re的不同步变化,致使在相同气温条件下,不同季节相同气温的NEE的差别较大。也说明了不同季节的气温高低对净生态系统碳收支的影响不同。
6 a日尺度上NEE随气温的变化趋势均是一样的,如表3所示,但每年的临界温度会有变化。在月尺度上,月总NEE对气温Ta的响应趋势与日尺度上的相似,也需要分段描述。这里用来分段的临界温度是通过2003—2008年6 a临界温度的平均值来设定,即T0=8.27 ℃。

表3 2003—2008年日尺度上Re对日平均5 cm处土壤温度Ts的响应趋势方程(y=aebx)系数与R2
通过分别分析在日尺度和月尺度上,长白山温带针阔叶混交林月尺度上NEE对气温(Ta)的响应趋势我们发现,NEE随气温的升高而逐渐向正的方向增大,但增大的幅度很小,都还在0附近,表现为碳排放;当到达一个临界温度时,NEE又随气温的升高而转变为向负的方向迅速增大,表现为碳吸收。同时还发现,每年的临界温度均有所不同,这说明在温度变化的影响下,生态系统也相应做出了调整,使得NEE发生转折的温度不同。
3 气候变化对该类生态系统碳收支对温度的响应特征的可能影响
基于月尺度生态系统碳通量,对比分析了2003—2008年长白山温带针阔混交林森林生态系统碳通量的年际变化特征中,每年季节尺度中碳交换各组分的最大值的变化以及其所对应的当月的月平均温度(气温和5 cm土壤温度)的变化特征,从而能够了解气候变化对生态系统碳收支各个组分的最适温度造成的影响。
2003—2008年每年月总GPP在7月达到最大值,因此可将7月的月平均气温和月平均5 cm土壤温度,分别看做是使GPP达到最大值的最适温度。表4是2003—2008年最大月总GPP与其相应月平均气温的对比(均出现在7月)。可以看出,每年的最适气温和最适5 cm土壤温度都是不同的,GPP也有相应的变化。GPP在2004、2007年出现了2个极小值,分别为288.5 gC·m-2·mon-1和301.7 gC·m-2·mon-1,根据最适气温和最适5 cm土壤温度的变化可以看出,2004、2007年出现的最适气温和最适5 cm土壤温度也相对较低,2004年的为18.8 ℃和16.2 ℃,2007年的为18.6和15.5 ℃。2008年的月总GPP达到最大值,为339.8 gC·m-2·mon-1,其对应的最适气温和最适5 cm土壤温度也相对较高,为20.1 ℃和16.3 ℃。这6 a的最适气温和最适5 cm土壤温度的平均值分别为19.1 ℃和16.3 ℃,每年的最适温度均分别在这2个温度值附近变化。总的来说,在不考虑其他影响因素的情况下,每年生长季节的最大GPP具有随其对应的最适温度升高而增大的趋势。
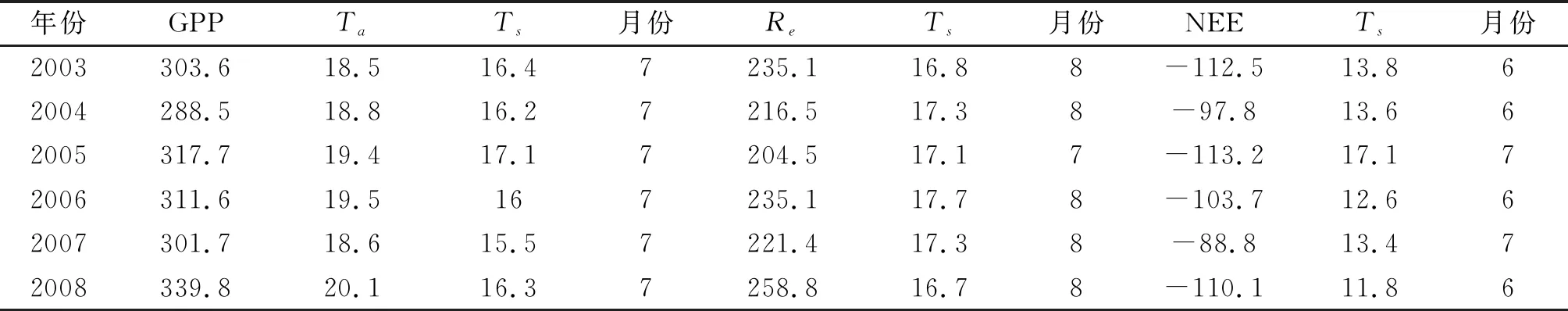
表4 2003—2008年长白山温带针阔混交林每年最大月总GPP、总Re,总NEE与其相应月月平均气温Ta和月平均5 cm土壤温度Ts的对比
2003—2008年月总Re与其相应月平均5 cm土壤温度的对比(表4)能够看出,2003—2008年生长季节出现最大月总Re的月份主要是在8月(除了2005年出现在7月),在2005年和2007年出现2个极小值,分别为204.5 gC·m-2·mon-1和221.4 gC·m-2·mon-1,所对应的土壤温度分别为17.1 ℃和17.3 ℃。这6 a平均每年Re所对应的最适5 cm土壤温度为17.1 ℃。由于长白山温带针阔混交林森林生态系统Re对5 cm土壤温度的敏感度更好,在这里主要考虑最适5 cm土壤温度的变化。可以得到,在季节尺度上,除了2003年和2008年外,其余年出现最高生态系统呼吸值随其相应的5 cm土壤温度升高而增大,随5 cm土壤温度的下降而减少。而2003年和2008年与其余年份不同可能是受这两年8月降水量较小等其他因素的影响。
由于NEE是GPP和Re之间的差值,而GPP和Re对温度的响应也有所不同。在季节尺度上,长白山温带针阔混交林生态系统出现最大月总GPP均在7月,而出现最大月总Re主要是在8月。由于时间上的不一致,NEE的最高值出现在GPP相对较高而Re相对较低的6月或7月。在这6 a中对于NEE的最适气温平均值为17.7 ℃。由表4可以看出,2005年NEE值负值方向最大为-113.2 gC·m-2·mon-1,其最适气温也较大为17.1 ℃。2004年、2007年NEE相对较小,其最适气温也较2005年低,在2003、2008年虽然最适气温较低,但是NEE负值方向较大,可能与降水、光照等其它环境因子有关。
根据2003—2008年月尺度上每年月总GPP、Re和NEE在数值上的最大值和其所对应的月平均气温和5 cm土壤温度(最适温度)的对比,得到每年的最适温度均有所变化。GPP在季节尺度上最大值出现在每年的7月,在不考虑其它影响因素的情况下,GPP随最适温度(气温和5 cm土壤温度)的升高而增大;在季节尺度上每年月总Re最大值主要出现在8月,在不考虑其他影响因素的情况下,最大月总Re值随其最适5 cm土壤温度升高而增大,随5 cm土壤温度的下降而减少。由于时间上的不一致,在季节尺度上每年的月总NEE的最高值出现在GPP相对较高而Re相对较低的6、7月。在2003—2008年间,由于2007年7月的月总GPP、Re值偏低,特别是7月的月总GPP值,使得2007年7月月总NEE在向正值方向上比其他几年大。通过生态系统碳收支各组分最大值及其相应的最适温度(气温或5 cm土壤温度)的变化,得到在气候变化的影响下,每年的最适温度也在变化,这使得每年的碳收支也在随之改变,但由于6 a的数据较少,变化趋势还不太明显,我们还需要更长时间的观测数据,开展更深入的分析。
4 结论
本文在通过分析长白山温带针阔混交林生态系统在2003—2008年年总GPP和Re在日尺度和月尺度上, GPP、Re随气温和5 cm处土壤温度变化均呈指数形式; NEE对气温的响应特征分2个阶段,即NEE随着气温升高而逐渐向正的方向增大,表现为碳排放,当到达临界气温后,NEE又随气温的迅速增长而转变为向负的方向的增大,当由正值转变为负值时,即由碳排放转变为碳吸收。在年尺度上,长白山针阔混交林生态系统碳收支主要受到温度和降水的影响。由于数据较少,该生态系统碳收支各组分与气温之间的关系不显著,我们在这里不做研究。该生态系统碳收支达到最大值时对应的最适温度(气温和5 cm土壤温度)在每年均有变化。这说明在气候变化的背景下,随着温度的变化,生态系统的最适温度也在改变。
由森林生态系统是具有复杂的结构和功能的有机体,不同时间尺度森林生态系统对温度变化的响应和适应机理存在显著的差异,对这些差异的理解和解释还需要更长时间的观测数据,开展更深入的分析[6]。
猜你喜欢
疯狂英语·新悦读(2022年7期)2022-11-23
今日中国·西班牙文版(2021年4期)2021-04-09
学生天地(2020年34期)2020-06-09
辽金历史与考古(2019年0期)2020-01-06
现代园艺(2018年2期)2018-03-15
中国环境科学(2016年4期)2016-05-27
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
安徽农业科学(2015年30期)2015-12-25
中国工程咨询(2015年8期)2015-02-16
