
植物多肽类胰蛋白酶抑制剂降糖功效研究进展
2021-11-14 11:24:48王雅馨石亚伟
食品工业科技 2021年22期
王 蕾,王雅馨,石亚伟
(山西大学生物技术研究所,化学生物学与分子工程教育部重点实验室,山西太原 030006)
胰蛋白酶抑制剂(Trypsin inhibitor,TI)通常是指一类具有抑制胰蛋白酶活性的物质,它能与胰腺分泌的丝氨酸蛋白酶系(如胰蛋白酶、胰凝乳蛋白酶等)发生不可逆的反应,进而降低生物体对蛋白质的消化、吸收,调节生物体内许多重要的生命活动。胰蛋白酶抑制剂来源广泛,普遍存在于动植物及微生物中。目前主要从豆科植物和动物肝脏中提取,在大多数植物贮藏器官中,其含量特别丰富,最高可达总蛋白的10%左右。迄今为止,已有数百种天然胰蛋白酶抑制剂相继得到分离和鉴定,并且有多种抑制剂被证明具有抗癌[1]、抗炎[2-3]、抗虫[4-5]及治疗心血管疾病[6]、人类获得性免疫缺陷症[7]等恶性疾病的潜在应用价值,相关的作用机制被广泛研究,但是对其在糖尿病治疗中的功效报道较少。
早在上世纪70年代,就有报道称大豆中微量的胰蛋白酶抑制剂对于糖尿病治疗、调节胰岛素失调有一定效果[8-10]。随后人们从大豆、银杏、永川豆豉、紫花芸豆、亚麻以及智利当地特有的一种豆科植物Geoffroea decorticansz中相继提取到了具有降糖活性的胰蛋白酶酶抑制剂,但其降糖功效仅在动物实验得以证实,缺乏相关降糖的分子靶标、作用机制等的研究。
本文将介绍胰蛋白酶抑制剂在降糖方面的作用及其研究现状,重点分析分子靶标是α-葡萄糖苷酶的降糖机制,并对胰蛋白酶抑制剂降糖功效的应用与开发提出展望,为今后深入研究胰蛋白酶抑制剂的降糖机理提供理论参考。
1 胰蛋白酶抑制剂的分类
胰蛋白酶抑制剂主要作用于分子内活性部位是赖氨酸的蛋白酶,属于丝氨酸蛋白酶抑制剂,能与胰蛋白酶(胰凝乳蛋白酶)的必需基团结合使其失活,从而抑制胰蛋白酶(胰凝乳蛋白酶)与底物结合。通常,人们根据胰蛋白酶抑制剂的相对分子质量、反应位点数以及半胱氨酸含量等差异,可将其分为五大主要类型:Kunitz型、Bowman-Birk型、Kazal型、PotatoⅠ及Potato Ⅱ型,其中对Kunitz、Bowman-Birk及PotatoⅠ/Ⅱ型的研究较多(见表1)。此外,还可根据其等电点的不同,将胰蛋白酶抑制剂分为酸性、中性、碱性这3种类型。
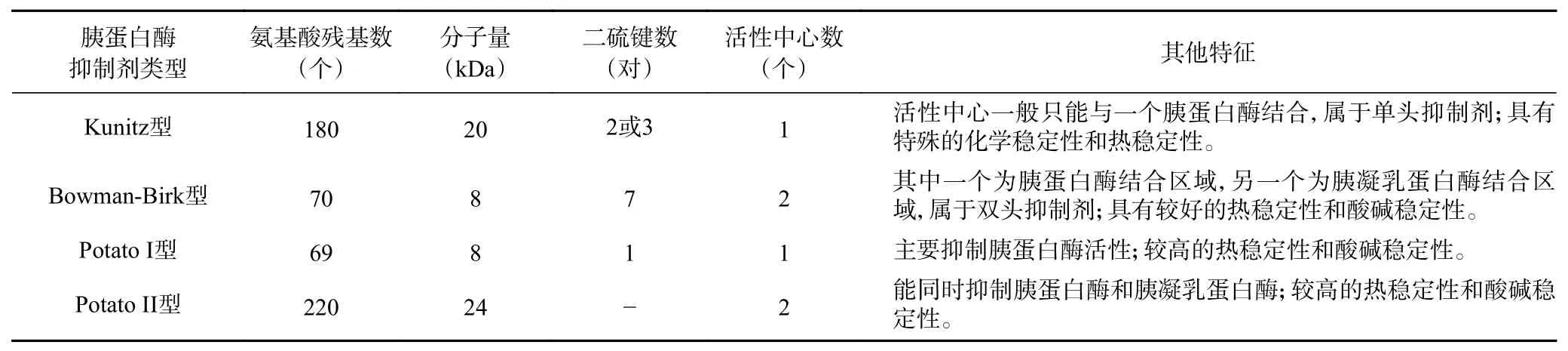
表1 胰蛋白酶抑制剂的分类及其特征Table 1 Classification and characteristics of trypsin inhibitors
1.1 Kunitz型胰蛋白酶抑制剂
Kunitz型胰蛋白酶抑制剂(Kunitz trypsin inhibitor,KTI)只含一个功能性抑制域和一个反应位点,只能抑制胰蛋白酶活性。KTI的三级结构较为保守,大多由一个明显的疏水核心、两对或三对高度保守的二硫键桥、三链β-折叠和一个N端310螺旋及一个C端α-螺旋组成(植物KTI的三维结构中不含有α-螺旋)[11-12]。
1.2 Bowman-Birk型胰蛋白酶抑制剂
Bowman-Birk型胰蛋白酶抑制剂(Bowman-Birk trypsin inhibitor,BBI)对胰凝乳蛋白酶、胰蛋白酶及弹性蛋白酶均可以发生抑制作用,不具有抑制专一性。BBI分子内富含半胱氨酸,可形成多个链内或链间二硫键,易折叠形成复杂的高级结构同时加上亚基间极性相互作用,使BBI具有较好的稳定性[13]。
1.3 Potato Ⅰ型和Potato Ⅱ型胰蛋白酶抑制剂
Potato Ⅰ型(PI-I)和Potato Ⅱ型(PI-II)胰蛋白酶抑制剂是一类具有较高热稳定性和酸碱稳定性的蛋白酶抑制剂。其中PI-I只有一个活性中心,主要抑制胰蛋白酶活性,而对胰凝乳蛋白酶的抑制作用较弱。PI-II由两个重复序列构成,包含两个活性中心,能同时抑制胰蛋白酶和胰凝乳蛋白酶的活性[14]。
2 胰蛋白酶抑制剂的降糖作用及其分子机制
虽然植物蛋白类胰蛋白酶抑制剂可能具有降糖活性早有报道,但对其降糖功效的研究近些年才被人们广泛关注。豆豉胰蛋白酶抑制剂可明显降低糖尿病小鼠血糖[15]。大豆胰蛋白酶抑制剂可以降低糖尿病模型小鼠血糖和血清甘油三酯水平,可改善其肝肾器官的病变程度[16],能够促进大肠对胰岛素的吸收[17]。紫花芸豆胰蛋白酶抑制剂可抑制四氧嘧啶诱导的糖尿病小鼠血糖升高,同时降低血清总胆固醇,但对高密度脂蛋白胆固醇和三酰甘油的效果不明显[18-19]。银杏种子中胰蛋白酶抑制剂可以有效的降低糖尿病模型大鼠血糖、血清甘油三酯、血清总胆固醇,同时升高高密度脂蛋白胆固醇[20]。花生中提取的一种胰蛋白酶抑制剂AHTI可以显著降低大鼠空腹血糖,同时能够控制体重增加和食物摄入量[21]。罗望子胰蛋白酶抑制剂TTI可以降低大鼠空腹血糖以及胰岛素抵抗指数,同时升高高密度脂蛋白胆固醇[22]。此外,亚麻胰蛋白酶抑制剂LUTI以及豆科植物Geoffroea decorticansz中的一种胰蛋白酶抑制剂GdTI也具有一定的降糖功效[23-25]。
2.1 具有降糖活性的胰蛋白酶抑制剂的类型
随着对胰蛋白酶抑制剂降糖作用研究的深入,研究人员发现其降糖功效并不具有普遍性。通过对目前有文献报道明确具有降糖活性且已知的胰蛋白酶抑制剂序列分析,可知亚麻胰蛋白酶抑制剂LUTI属于Potato I型胰蛋白酶抑制剂,pI值为9.55[23];大豆胰蛋白酶抑制剂SBTI和罗望子胰蛋白酶抑制剂TTI属于Kunitz型胰蛋白酶抑制剂,其pI值分别为8.37和5.41[16,22](见图1)。而针对近期报道的一种豆科植物Geoffroea decorticansz中的胰蛋白酶抑制剂GdTI的研究发现,其已知的一段酶解序列与大豆中的BBI同源性较高,但没有完整的氨基酸序列信息,属于Bowman-Birk型胰蛋白酶抑制剂的类型。从目前文献资料来看,各种类型的胰蛋白酶抑制剂均有可能具有降糖作用,其共性在于具有胰蛋白酶的抑制活性,但如何发挥其在动物体内的降糖作用,从一级结构序列很难看出其他共同特点(序列比对使用Genetyx和GeneDoc软件完成)。

图1 LUTI, SBTI和TTI的同源序列比对Fig.1 Alignment of the amino acid sequence of LUTI, SBTI and TTI
2.2 胰蛋白酶抑制剂降糖作用的分子机制
目前,已知报道的降糖药物作用机制如图2所示,主要包括:肠道内微生物菌群的调控及延缓肠道内葡萄糖的吸收(糖苷酶抑制剂等)[26-27];通过提高PPARγ表达量和AMPK活性等增加靶细胞对胰岛素的敏感性(胰岛素增敏剂等)[28-29];促进胰岛β细胞分泌胰岛素(磺脲类、格列奈、GLP-1类似物等)[30];增强葡萄糖转运蛋白等的表达以促进葡萄糖吸收,增加外周组织摄取和利用葡萄糖(双胍类)[31]。此外,还可以通过降低炎症因子TNF-α、IL-4、IL-17等表达量减少炎症反应以降低线粒体氧化应激损伤[32-33],提高胰岛素与受体的亲和力以增加胰岛素对血糖的清除作用[34],降低胰岛β细胞及胰岛组织损伤[35],抑制肝糖异生和糖原分解、降低肝糖输出、减少糖原生成[36]等途径来达到降糖的目的。
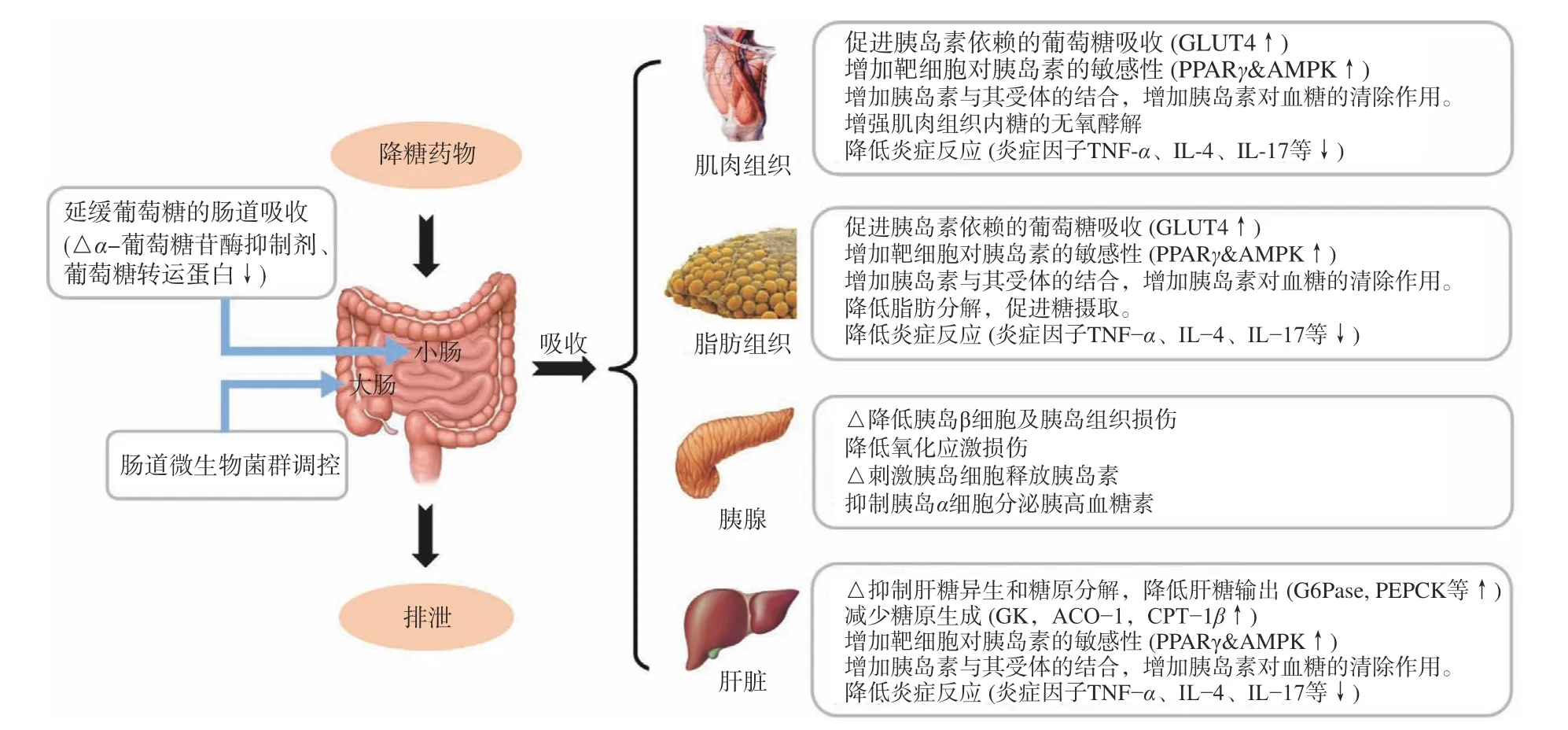
图2 体内改善葡萄糖稳态的相关机制汇总Fig.2 Summary of potential mechanisms to improved glucose homeostasis
胰蛋白酶抑制剂虽已经证实具有降糖功效,但是其具体的降糖方式、作用靶标并不十分清楚。目前研究较为集中的分子靶标是α-葡萄糖苷酶,通过抑制其活性来达到降糖的功效,此外在抑制糖异生、促进胰岛素分泌、免疫调控等方面也有一定的报道,可以说胰蛋白酶抑制剂的降糖功效可能是一种或多种机制共同作用的效果(如表2)。

表2 胰蛋白酶抑制剂的降糖机制Table 2 Hypoglycemic mechanism of trypsin inhibitors which has been reported
2.2.1 分子靶标是α-葡萄糖苷酶的胰蛋白酶抑制剂
在以往降糖药物的研究中,α-葡萄糖苷酶抑制剂一直都是降低餐后血糖的一线药物,其能够通过竞争性抑制或可逆性抑制位于小肠刷状细胞表面的α-葡萄糖苷酶活性、延缓总体碳水化合物的消化时间、调节餐后胃肠激素和肠道菌群等方式有效控制餐后血糖的升高,产生较为理想的降糖效果[38]。天然植物是α-葡萄糖苷酶抑制剂的主要来源,但是目前绝大多数研究主要集中在黄酮类[39-40]、皂苷类[41]、三萜类[42]、奎宁酸类[43]、酚类[44]、苯乙胺类[45]等化合物中,而对于植物蛋白类的研究较少。Ye等[46]从白色扁豆中分离得到一种28 kDa的蛋白dolichin,通过抑制α-葡萄糖苷酶的活性来调节血糖水平。大花田青中分离的蛋白SGF60/90以及豇豆中的一种抗病毒蛋白unguilin对α-葡萄糖苷酶具有显著的抑制作用[47-48]。此外,人们还从大麻种子[49]、苦瓜[50]、白蛋白[51]、南瓜子[52]、大豆[53]、牛奶[54]等提取了不同种类的降糖多肽,研究表明其调节血糖的分子靶标正是α-葡萄糖苷酶。近期,Wang等[24]从胡麻的籽粕分离到一个分子量为8 kDa的小分子多肽,经质谱鉴定该多肽为亚麻胰蛋白酶抑制剂LUTI,其可与α-葡萄糖苷酶在体外紧密结合,竞争性抑制α-葡萄糖苷酶活性,其IC50值为113.92 μmol/L(阿卡波糖为592.35 μmol/L),且在细胞水平上能够增加葡萄糖的消耗和乳酸的生成,从而达到降糖的功效。Cotabarren等[25]从智利当地特有的一类豆科植物Geoffroea decorticansz中分离得到一种胰蛋白酶抑制剂GdTI,对α-葡萄糖苷酶具有很强的抑制活性(IC50=0.18 μmol/L),同时该抑制剂具有良好的抗凝血功能,这为其作为调控血糖的潜在药物提供了一定的可能性。
此外,研究人员在以α-葡萄糖苷酶为分子靶标的研究中发现,胡萝卜中存在一种碱性胰蛋白酶抑制剂DCTI能够抑制α-葡萄糖苷酶活性(其半抑制常数稍低于GdTI),但是其并不具有胰蛋白酶抑制活性(未发表数据)。通过序列分析可知这种胰蛋白酶抑制剂属于Potato I型胰蛋白酶抑制剂,且与LUTI的同源性为69.84%(如图3)。但同属Potato I型的甜荞麦胰蛋白酶抑制剂BTI(Fagopyrum esculentum,common buckwheat)则不具有α-葡萄糖苷酶抑制活性[24]。

图3 Potato I型胰蛋白酶抑制剂的同源序列比对Fig.3 Alignment of the amino acid sequence of Potato I trypsin inhibitors
2.2.2 分子靶标是非α-葡萄糖苷酶的胰蛋白酶抑制剂 胰蛋白酶抑制剂的降糖机制,除通过抑制α-葡萄糖苷酶活性外,其是否会影响到其他的降糖靶标(如葡萄糖转运蛋白、胰岛素分泌、胰岛素敏感性、胰岛素抵抗、炎症反应、糖代谢等)还不十分清楚(如图2),人们对于胰蛋白酶抑制剂降糖作用机理的认识还比较有限。豆豉胰蛋白酶抑制剂对四氧嘧啶糖尿病模型小鼠胰岛组织有明显的修复作用,推测其可能是通过保护或修复胰岛β细胞及胰岛组织,促进胰岛素分泌或释放从而达到降糖的目的,但其对α-葡萄糖苷酶无抑制活性,同时也不影响糖代谢过程[15]。王静等[18]推测紫花芸豆胰蛋白酶抑制剂的降糖机制可能是通过其对胰腺产生作用,促进胰岛素分泌释放,从而增加各个组织对葡萄糖的摄取和利用。而对大豆胰蛋白酶抑制剂SBTI的研究发现,其可以保护四氧嘧啶糖尿病模型小鼠的肝细胞、改善其糖耐量、抑制糖异生,促进肝糖原合成,进而使血糖浓度下降。陈金铭等[20]发现银杏种子中胰蛋白酶抑制剂可以通过有效的降低血清中细胞免疫因子TNFα、IL-4、IL-17和免疫球蛋白IgA水平,升高高密度脂蛋白胆固醇,免疫球蛋白IgG、IgM水平,对免疫系统及补体系统有一定的免疫调节作用,达到治疗糖尿病目的。Serquiz等[21]发现花生胰蛋白酶抑制剂AHTI能够增加饱腹感,促进人胆囊收缩素/肠促胰酶肽(CCK)的生成,促进胰岛素分泌,进而降低空腹血糖。罗望子胰蛋白酶抑制剂TTI很可能通过抑制脂肪组织分泌的激素作用从而降低空腹血糖,或者作为一种激素信号与胰岛素受体结合,参与葡萄糖的吸收和释放[22,55-56]。
3 展望
胰蛋白酶抑制剂通常被认为是一种抗营养因子,在动物养殖及人类食品的应用中都需要经特殊处理使其失活,才能有助营养的吸收和利用。由于天然植物来源的胰蛋白酶抑制剂存在范围广、稳定性好,与一般小分子药物相比毒副作用小、开发成本低且对消化道内的蛋白酶具有天然的抵抗能力等优点,其在降糖功能性食品开发以及口服药物糖尿病的预防、治疗中有巨大的潜在效用,因此,开展对不同类型胰蛋白酶抑制剂的降糖活性及其降糖机理的研究就显得尤为重要。一直以来,人们对胰蛋白酶抑制剂的研究主要聚焦于抗病毒、抗癌等多种功效上,而对其降糖功效的研究虽在体外动物实验得以证实,但其具体的降糖效果及降糖机制还不明确,关于胰蛋白酶抑制剂是否能作为一种药品治疗糖尿病,或是作为一种功能性食品、保健品预防糖尿病的研究还需要继续深入。未来研究需要侧重在以下两方面:一是胰蛋白酶抑制剂的降糖功效是否具有普遍性,从结构和某些生化特性上明确具有降糖活性的胰蛋白酶抑制剂类型,为找寻更多的此类药物提供某种可能性;二是胰蛋白酶抑制剂的降糖机制具体是什么,通过对其降糖机理的挖掘,可以进一步运用生物化学、分子生物学等手段提高其降糖效果,减少药物的不良反应,对于应用其防治糖尿病和提高患者生存质量方面具有重要的现实意义。
猜你喜欢
军民两用技术与产品(2021年10期)2021-03-16 06:05:10
基层中医药(2020年6期)2020-09-11 06:35:28
世界农药(2019年3期)2019-09-10 07:04:10
中成药(2018年12期)2018-12-29 12:25:38
中成药(2018年6期)2018-07-11 03:01:14
中成药(2018年5期)2018-06-06 03:11:58
基层中医药(2018年1期)2018-03-01 07:36:13
中成药(2017年8期)2017-11-22 03:19:32
光学精密工程(2016年4期)2016-11-07 09:04:48
肿瘤影像学(2015年3期)2015-12-09 02:38:45
