
不同林龄杉木人工林的林下植被与土壤理化特性
2021-11-11 09:00:48张利荣李惠通郑立津陈丽敏刘雨辉刘爱琴
亚热带农业研究 2021年3期
张利荣, 李惠通, 郑立津, 陈丽敏, 刘雨辉, 刘爱琴
(1.福建农林大学林学院,福建 福州 350002; 2.国家林业局杉木工程技术研究中心,福建 福州 350002;3.福建农林大学莘口教学林场,福建 三明 365000)
林下植被是森林生态系统的重要组成部分,对维护森林生态系统的稳定性和恢复力、保持土壤肥力、提高林地生产力具有重要作用[1]。土壤是植被生长的基质,在驱动植物群落演替上起关键作用。植物群落会随着土壤养分种类及含量的变化而发生转变,从而改变植物物种多样性[2]。而不同种类的植物对土壤中营养元素吸收量的不同也会导致土壤养分含量发生变化,从而影响土壤的稳定性[3]。杉木[Cunninghamialanceolata(Lamb.) Hook]作为我国南方特有的速生丰产常绿针叶树种,具有干材通直、材质佳、产量高和易种植等优点,在我国南方17个省(区)广泛种植[4-6]。根据第九次森林资源清查数据,目前我国杉木人工林面积已达986.7万hm2,蓄积量达到7.55亿m3,居全国人工林总面积及总蓄积量首位[7]。随着人工林面积的增加,地力衰退问题日益严重,有关人工林林下植被与土壤因子的关系越来越受关注。陈春如等[8]、张勇强等[9]探讨了不同密度下杉木人工林林下植被多样性;赵苏亚等[10]、李萌等[11]研究表明,间伐强度可以改变不同功能类群的丰富度,如轻度间伐可以提高木本植物功能类群的丰富度,而重度间伐则提高蕨类植物的丰富度;姜俊等[12]研究发现,近自然经营改造下杉木人工林林下灌木层和草本层的丰富度指数均高于未改造纯林。本文以近自然经营的不同林龄杉木人工林为研究对象,比较其林下植被多样性,并探究林下植被多样性与土壤肥力间的相关性,以期为杉木人工林地力维护提供参考。
1 材料与方法
1.1 研究区概况
试验林地设置在福建省三明市福建农林大学莘口教学林场(117°25′~117°32′E,26°7′~26°12′N),属亚热带季风气候,地貌为低山丘陵,年均气温约19.1 ℃、年均降雨量约1 749 mm,土壤为红壤土。试验地均为二代改良杉木人工林,造林前经炼山整地,林下植被灌木层主要有山苍子(Litseacubeba)、楤木(Araliachinensis)、东南野桐(Mallotuslianus);草本层主要为枇杷叶紫株(Callicarpakochiana)、乌毛蕨(Lechnumorientale)、五节芒(Miscanthusfloridulus)等。样地基本情况详见表1。

表1 杉木人工林样地概况Table 1 Overview of Chinese fir plantation sites
1.2 样地设置及调查方法
2017年4月选择经营条件、立地质量和坡度基本一致的幼林龄(5年生)、中林龄(15年生)和成熟林(24年生)独立林分。每个林分选择5个大小为20 m×20 m的代表性样地,样地上下边界均平行于等高线,四角分别设置带有编号的水泥桩,共设15个标准样地。每个样地按“S”形(梅花形)设置5个2 m×2 m的小样方,对样方内胸径小于4 cm的乔木和灌木进行统计调查,记录其株数、地径、高度以及冠幅等;每个样地按“S”形设置5个1 m×1 m小样方,对林下草本、蕨类、藤本植物进行统计调查,记录各层植物的种类、个数、高度和盖度等。
每个样地以“S”形选取5个点,分别挖宽×高为50 cm×30 cm的垂直剖面。用环刀(100 cm3)在剖面左侧0~20 cm内取原状土样,用于测定土壤水分和物理性质;取剖面右侧0~20 cm内土样,并将同一样地内的土壤进行等量充分的混合,随机选取混合土样1 kg存于自封袋中,并带回实验室。将混合土样自然风干后研磨,并过2 mm尼龙土壤筛,再以四分法取100 g风干土样过0.149 mm尼龙土壤筛,分别装入自封袋,用于土壤养分的测定。其中,土壤总氮含量用碳氮元素分析仪(德国Elementar)测定;有机质含量采用重铬酸钾外加热法测定;pH值采用电位法(土∶水=1∶2.5)测定;全磷和有效磷含量选用钼锑抗比色法测定;全钾含量用火焰光度法测定[13]。
1.3 统计与分析

采用SPSS 20.0软件的单因素方差分析(ANOVA)检验,分析不同样地土壤理化性质的差异性及其林下植被与土壤间的相关性。
2 结果与分析
2.1 不同林龄杉木人工林林下植被物种组成
参试杉木人工林林下植被灌木层和草本层组成见表2。由表2可知,幼龄林林下植被共有42科52属60种,中龄林有41科51属59种,成熟林有38科48属54种。由表2还可知,灌木层植被科属种数量均随着林龄的增加而减少,而草本层植被中仅种的数量随林龄的增加呈递增趋势。分析其原因,可能是随着杉木林龄的增加,林分郁闭度增大,林木间竞争力增强,林下植被生长环境及光照条件发生改变,喜光的灌木生长优势减弱,阴生植物逐渐凸显。

表2 不同林龄杉木人工林林下植被组成Table 2 Composition of undergrowth vegetations of Chinese fir plantations of different forest ages
2.2 不同林龄杉木人工林植物的重要值
2.2.1 乔木层 由表3可知,不同林龄杉木人工林乔木种类排序依次为:中龄林(8种)>幼龄林(7种)>成熟林(6种)。幼龄林乔木层内重要值排前三的是杉木、木荷(Schimasuperba)和毛叶冬青(Ilexmicrococca);中龄林排前三的是杉木、毛叶冬青和木荷;成熟林排前三的有杉木、绒毛润楠(Machilusvelutina)和细齿叶柃(Euryanitida)。杉木在不同林龄中均为优势种,其重要值分别为14.72、17.05和18.27。不同林龄杉木人工林乔木层有3种共有种,分别为杉木、绒毛润楠和吊皮锥(Castanopsiskawakamii)。其他种类的乔木出现频度较低,群落作用不明显且分布较散,为群落中的偶见种。

表3 不同林龄杉木人工林植物物种组成特征Table 3 Plant species composition characteristics of undergrowth vegetation of Chinese fir plantations of different ages

续表3
2.2.2 灌木层 不同林龄杉木人工林林下灌木种类排序依次为:成熟林(15种)>中龄林(14种)>幼龄林(13种)。幼龄林灌木层内重要值排前三的是枇杷叶紫珠、长圆叶鼠刺(Iteachinensis)和粗叶榕(Ficushirta),重要值分别为111.81、19.09和9.93;中龄林排前三的是箬竹(Indocalamustessellatus)、粗叶榕和枇杷叶紫珠;成熟林排前三的有箬竹、粗叶榕和山血丹(Ardisialindleyana)。中龄林和成熟林的优势种均为箬竹,重要值分别为42.74和66.13。不同林龄杉木人工林灌木层共有种包括:长圆叶鼠刺、玉叶金花(Mussaendapubescens)、山血丹、粗叶榕、箬竹、锈毛莓(Rubusreflexus)。不同林龄林分灌木层均发现偶见种,但频率不高,如幼龄林中的变叶榕(Ficusvariolosa)和山油麻(Tremadielsiana),中龄林中的柏拉木(Blastuscochinchinensis)和短梗新木姜子(Neolitseabrevipes),成熟林中的黄花倒水莲(Polygalaaureocauda)和山杜英(Elaeocarpussylvestris)等。经比较,成熟林中的灌木层稀有种相对较多。
2.2.3 藤本植物 不同林龄杉木人工林藤本植物种类排序依次为:幼龄林(8种)=中龄林(8种)>成熟林(7种)。幼龄林藤本植物重要值排前三的是细圆藤(Peicampylusglaucus)、异果崖豆藤(Millettiadielsiana)和海金沙(Lygodiumjaponicum);中龄林排前三的是细圆藤、海金沙和短梗南蛇藤(Celastrusrosthornianus);成熟林排前三的是细圆藤、毛鳞省藤(Calamusthysanolepis)和网脉酸藤子(Embeliarudis)。细圆藤在不同林龄林分中均为优势种,重要值分别为8.55、16.77和17.71。不同林龄林分藤本植物共有种较为一致,相比于成熟林的稳定,幼龄林和中龄林的物种出现频率较低。
2.2.4 草本层 不同林龄杉木人工林草本种类排序依次为:幼龄林(11种)>中龄林(8种)=成熟林(8种)。幼龄林草本层重要值排前三的是五节芒、棕叶狗尾草(Setariapalmifolia)和黑莎草(Gahniatristis),重要值分别为83.64、41.13、16.72;中龄林排前三的是金毛狗(Cibotiumbarometz)、华山姜(Alpiniachinensis)和棕叶狗尾草;成熟林排前三的是金毛狗、华山姜和深绿卷柏(Selaginelladoederleinii)。中龄林和成熟林的优势种和次优势种一致。根据出现频率与数量分析发现,幼龄林中的华麻花头(Serratulachinensis)和射干(Belamcandachinensis)以及铜锤玉带草(Pratianummularia)属于偶见种,且未出现在中龄林和成熟林中,是杉木林中的稀有种。
2.2.5 蕨类植物 蕨类丰富度随着林龄的增加而增大,成熟林蕨类植物种类多于中龄林和幼龄林,可能与蕨类植物喜湿润的环境有关。成熟林郁闭度较高,使得林下湿度较高,较适宜蕨类植物生长。3种林龄杉木人工林蕨类共有种为狗脊蕨(Cibotiumbarometz)、扇叶铁线蕨(Serratulachinensis)、羽裂圣蕨(Dictyoclinewilfordii)和乌毛蕨(Lechnumorientale)。幼龄林和中龄林的优势种是乌毛蕨,重要值分别为103.39和71.28;次优势种为狗脊蕨,重要值分别为36.03和66.17。成熟林的优势种为扇叶铁线蕨,重要值为68.12;次优势种为淡绿短肠蕨(Allantodiavirescens),重要值为61.81。
2.3 不同林龄杉木人工林的多样性指数
由表4可知,成熟林灌木层丰富度指数小于中龄林和幼龄林,但各林分间差异不显著。中龄林草本层丰富度指数显著大于幼龄林和成熟林。灌木层和草本层优势度指数随着林龄的增加呈先升高后降低的趋势,其中灌木层中龄林优势度指数显著大于成熟林,草本层中龄林显著大于幼林龄。灌木层多样性指数各处理间差异不显著;草本层多样性指数则呈先上升后下降的趋势,且中龄林显著大于幼龄林。灌木层均匀度指数随林龄的增加呈上升趋势,成熟林灌木层均匀度指数大于幼林龄和中林龄;草本层均匀度指数随林龄的增加呈先上升后下降的趋势,且中龄林和成熟林显著大于幼龄林。总体来看,林龄对灌木层丰富度差异不显著;中龄林草本层各多样性指数较幼龄林和成熟林均有所提升,其中丰富度指数显著大于幼龄林和成熟林,优势度指数、多样性指数和均匀度指数均显著大于幼龄林。

表4 不同林龄杉木人工林的多样性指数1)Table 4 Diversity index of undergrowth vegetation of Chinese fir plantations of different forest ages
2.4 不同林龄杉木人工林林下植被多样性与土壤理化性质的相关性
2.4.1 土壤理化性质 由表5可以发现,不同林龄杉木人工林土壤最大持水量、田间持水量、总孔隙度、质量含水量均随林龄的增加呈先下降后上升的趋势,容重随林龄的增加呈现先上升后降低的趋势,毛管孔隙度则随林龄的增加而升高。不同林龄的田间持水量、总孔隙度、毛管孔隙度、质量含水量均无显著差异;幼龄林的最大持水量显著大于中龄林和成熟林,但容重显著小于中龄林和成熟林。

表5 杉木人工林样地土壤理化性质(0~20 cm)1)Table 5 Soil physical and chemical properties of Chinese fir plantation sites(0~20 cm)
土壤有机质、全磷含量及pH值随着林龄的增加呈先降低后升高的趋势,全氮含量则呈现上升的趋势,有效磷和全钾含量呈先升高后降低趋势。不同林龄间有机质、有效磷含量无显著差异; 成熟林全氮、全磷含量显著高于中龄林和幼龄林;幼龄林全钾含量显著低于中龄林和成熟林。幼龄林pH值高于中龄林和成熟林。
2.4.2 物种多样性指数与土壤理化性质的相关性 草本层和灌木层物种多样性指数与土壤环境因子的Pearson 相关分析表明(表6),总孔隙度、毛管孔隙度、质量含水量及有机质、全氮、全磷、有效磷含量与各多样性指数不存在明显的相关性。最大持水量、田间持水量与草本层均匀度指数呈显著负相关,与其他多样性指数相关性不显著;土壤容重与草本层生态优势度指数呈显著正相关,与其他多样性指数相关性显著;全钾与草本层物种多样性指数呈显著正相关,与其他多样性指数相关性不显著。土壤pH与草本层生态优势度指数、多样性指数呈显著负相关,与其他多样性指数无显著相关性。
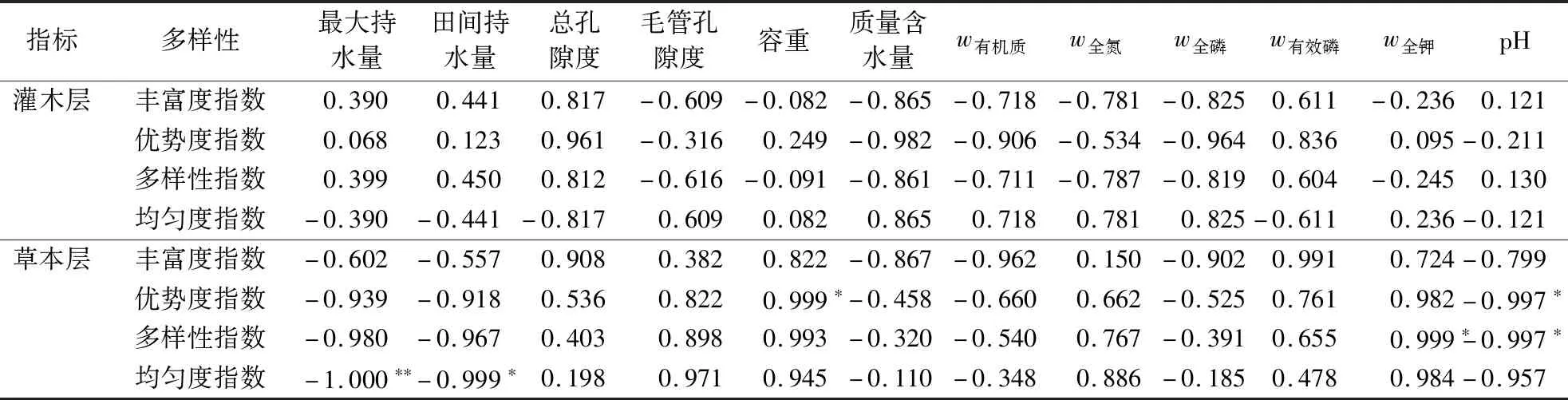
表6 物种多样性指数与土壤理化性质的相关关系分析1)Table 6 Analysis of correlation between species diversity index and soil factors
3 讨论与结论
物种多样性反映了生物群落在组成、结构、功能和动态方面的异质性,体现了群落结构类型、组织水平、发展阶段、稳定程度和生境的差异[16]。本研究表明,灌木层丰富度指数、优势度指数、多样性指数、均匀度指数随林龄增加呈先升高后降低的趋势。王敏等[17]研究表明,四川威远不同林龄桉树人工林灌木层物种分布、组成以及物种多样性随林龄的增加呈先升高后降低的趋势,与本研究结果一致。而薛云展等[18]研究湖南芦头不同林龄杉木人工林的物种多样性发现,随着林龄的增大物种多样性呈减小趋势,分析原因可能与芦头森林生态站森林健康等级处于亚健康状态,而且林分密度过高导致林分内宿存枯枝、枯死木较多[19],林分结构变差,透射光、散射光、通风程度等降低有关。本研究还发现,中龄林和成熟林的植物均匀度普遍高于幼龄林,灌木层物种多样性指数高于草本层,但草本层的物种多样性指数和均匀度指数波动幅度较大,分析其原因可能是林地单位面积内保留活立木株数不同,使得林分内光照有明显的时间和空间差异[6],光照增加使林分透光度增强,喜光的灌木层植物竞争增强。相对于草本层,灌木层对杉木人工林林下生境的适应能力更强,而草本层植物对环境的变化更为敏感,生长周期短且有随机性[20]。
林下物种群落的演替和更新进程会改变土壤理化性质,而土壤水分和养分的变化也会影响林下植物群落的演替。因此,提高土壤稳定性可为植物的定居、生存以及生长发育提供良好的生活环境[21]。本研究发现,不同林龄土壤的理化指标不同,成熟林土壤有机质、质量含水量、全氮及全磷含量相比幼林龄和中林龄均有所升高,说明杉木人工林成熟林的土壤肥力有所提升,地力衰退得到适当的缓解,这与崔宁洁等[22]的研究结果相同。可能是因为成熟林林分郁闭度适合林下植被接受充足的光照,形成更好的林内水热条件,枯落物分解速度增快,有效缓解了因林木砍伐造成的养分输入量减少[23-24];同时林下植被的凋落物积累,为土壤提供了丰富的有机质输入源,故土壤肥力水平较高[25]。群落演替过程中物种多样性变化与土壤因子存在着一定的相关性,其相互作用在改善土壤养分和促进物种多样性及其恢复等方面起着关键作用[26]。相关分析发现,草本层均匀度指数与田间持水量、最大持水量呈负相关性;草本层多样性指数与土壤pH呈负相关,与全钾含量呈正相关;草本层优势度指数与土壤容重呈正相关,与土壤pH呈负相关。这与盛茂银等[26]研究植被多样性与土壤因子相关性的结果不尽相同,可能与调查方法、树种、气候以及立地质量的差异有关。关于林下植被与土壤因子的相关性研究还有待进一步深化。
猜你喜欢
乡村科技(2023年20期)2024-01-08 07:20:00
江西农业大学学报(2022年1期)2022-02-04 11:51:42
江西农业大学学报(2021年2期)2021-05-16 02:18:24
农业与技术(2018年16期)2018-11-28 11:24:08
森林工程(2018年5期)2018-05-14 13:54:30
现代农业科技(2017年10期)2017-07-12 14:14:49
防护林科技(2016年5期)2016-09-05 01:19:50
广西林业科学(2016年2期)2016-03-20 05:53:41
中国农业信息(2016年17期)2016-02-06 09:50:05
农业与技术(2014年9期)2014-10-20 08:57:37
