
Relationship between TERT Polymorphism and Telomere Length in Workers Exposed to Omethoate*
2021-11-11 12:39:54CHENGShuaiLIUBinGUOZhiFengDUANXiaoRanLIUSuXiangLILeiYAOWuYANGYongLiandWANGWei
CHENG Shuai, LIU Bin, GUO Zhi Feng, DUAN Xiao Ran, LIU Su Xiang, LI Lei,YAO Wu, YANG Yong Li, and WANG Wei,4,#
Omethoate is a highly toxic organophosphorus pesticide that is widely used in agricultural production because of its high efficiency, broad spectrum, and low residue. Organophosphorus pesticides, such as omethoate, can inhibit acetylcholinesterase activity, leading to the accumulation of the neurotransmitter acetylcholine at cholinergic synapses. Accumulation of acetylcholine continues to stimulate cholinergic receptors, causing central nervous effects. Studies have shown that organophosphorus pesticides can cause genotoxicity in a variety of organisms,resulting in chromosomal DNA damage[1].
Telomeres are DNA-protein structures consisting of tandem hexamer repeats (TTAGGGn) at the end of the chromosome. They play important roles in chromosomal location, replication, protection, and control of cell growth. Telomerase is a DNA polymerase that synthesizes the TTAGG sequence and uses internal RNA molecules as templates to lengthen the pre-existing 3'terminal telomeres in vertebrates. It is a complex system composed of telomerase reverse transcriptase (TERT), telomerase RNA component and telomerase-associated protein 1 (TEP1).TERTis an important factor in maintaining telomere DNA length and chromosome stability.TERTis silently expressed in normal somatic and non-proliferative cells. However, in many human cancers, theTERTpromoter mutates, resulting in abnormal expression.TEP1, another component of the telomerase nucleoprotein complex, catalyzes the addition of new telomeres to chromosomes.TEP1immunoprecipitation has revealed its telomerase activity and its relatedness toTERTand telomerase RNA components[2].
A study of American adults found that the environmental exposure levels to organophosphate pesticides is related to alteration in telomere length in the population[3]. We have previously studied the relationship between tankyrase (TNKS) gene polymorphism and telomere length in peripheral blood leukocytes. The results showed that the CG+CC genotypes in rs1055328 may affect omethoate-induced telomere length increase[4].Additionally, in a study on the relationship between metabolizing enzyme gene polymorphisms and telomere length in omethoate-exposed workers, the extension of telomere length was related to glutathione S-transferase M1 (GSTM1) deletion,GG+AG genotypes, and interactions between smoking and GG+AG genotypes[5]. However, it is not clear whether the effect of omethoate on telomere length is related to the polymorphism of telomerase genes. Therefore, we investigated the effect of polymorphisms inTERTandTEP1on the telomere length of workers exposed to omethoate.
A total of 180 workers exposed to omethoate for more than 8 years were selected as the exposure group and 115 healthy people who were not exposed to toxic substances were selected as the control group. Smokers were defined as those smoking more than one cigarette a day for over half a year; alcohol drinkers were defined as those drinking more than twice a week in the past half year. This study was approved by the Life Science and Ethics Review Committee. Ten polymorphic loci associated with these genes (TERT: rs2736109,rs2735940, rs3215401, and rs2736100;TEP1:rs1713449, rs1760897, rs1760903, rs938886,rs1760904, and rs4246977) were studied by NCBISNP or Hapmap databases. The AssayDesigner3.1 software was used to design PCRs and single-base extension primers. The primer sequences of each of the polymorphic loci are listed in Supplementary Table S1, available in www.besjournal.com. Realtime fluorescence quantitative PCR assay was used to detect the DNA telomere length in peripheral blood leukocytes, and each sample was tested twice.Telomere length was determined using reference and telomere primers. The reverse and forward primers for the reference gene werehbgd,5'-GCCCGGCCCGCCGCGCCCGTCCCGCCGGAGGAGAAGTCTGCCGTT-3' andhbgu, 5'-CGGCGGCGGGCGGCGCGGGCTGGGCGGCTTCATCCACGTTCACCTTG-3'. The reverse and forward primers for the telomere were 5'-TGTTAGGTATCCCTATCCCTATCCCTATCCCTATCCCTAACA-3' and 5'-ACACTAAGGTTTGGGTTTGGGTTTGGGTTTGGGTTAGTGT-3'.
Statistical software (SPSS 21.0) was used for data analysis. In this study, data on telomere length were non-normally distributed. The telomere length data of the exposed and control groups were converted to normal distribution data using the Ln(X)+3 logarithmic transformation method. Thet-test was used to compare the differences in telomere length between the two groups. The covariance method was used to analyze the relationship between variables and telomere length. The factors influencing telomere length in omethoate workers were analyzed using generalized linear models. All statistical tests were two-sided, with a statistical significance level of α = 0.05.
The telomere length in the exposed group (3.52 ±0.62) was longer than that in the controls (3.00 ±0.36) (t= 9.108,P< 0.001). Additionally, we analyzed the effects of sex, age, smoking, alcohol consumption, and omethoate exposure on telomere length. The results showed that, except in smokers,the telomere length in the exposed group was significantly longer than that in the control group(P< 0.05) (Supplementary Table S2, available in www.besjournal.com). After the Hardy-Weinberg equilibrium test, the genotype distribution of each genetic polymorphism did not deviate (P> 0.05),indicating that the control group was representative.Covariance analysis was used to analyze differences in telomere length between different genotypes of theTERTandTEP1polymorphisms (Table 1). At theTERTrs2736109 polymorphism, the telomere length of the GG genotype was close to that of the AG genotype, thus leading to their fusion. The results showed that the telomere length of the AG+GG genotype was significantly longer than that of the AA genotype in the exposed group (P= 0.029). In the control group, the telomere length of the TT genotype of theTERTrs2736100 polymorphism was shorter than that of the GT genotype (P= 0.037).There were no significant differences in genotypes between the other loci. The rs2736100 genetic variation is associated with a range of cancers and related disorders. A case-control study of 828 people suggested that individuals with TG or GG had a higher risk of non-small-cell lung cancer compared to individuals with TT in the rs2736100 genotype[6]. Gu et al.[7]reported that the G allele of rs2736100 is significantly associated with a decrease in telomere length in Caucasians. However, we found that the telomere length of theTERTrs2736100 TT genotype was significantly lower than that of the GT genotype in the normal control population (P= 0.037), this could be due to ethnic differences.
In the generalized linear model, telomere length was used as the dependent variable; exposure,TERTrs2736109, andTERTrs2736100 were used as independent variables; and sex, age, smoking,drinking, and working period were used as covariates to enter the model. Generalized linear model analysis showed that exposure (b = 0.568,P<0.001) andTERTrs2736109 (GG+AG) (b = 0.240,P=0.045) affected telomere length, and no other factors were found to affect telomere length(Table 2). The polymorphism rs2736109 is located in the specific promoter region ofTERT. The specific binding of theTERTpromoter and the transcription factor GATA-2 can initiate theTERTtranscription process. All members of the GATA transcription factor family bind to a specific nucleotide sequence(T/A (GATA) A/G). Instead of the G-allele inTERTpromoter, the mutant A allele in rs2736109 generates a new GATA-1 binding locus[8], which decreases the transcription efficiency ofTERTby competitive inhibition. GATA-1 encodes two zinc finger structure motifs, c-terminal zinc finger (c-znf)and N-terminal zinc finger (n-znf). N-znf, interacts with the nuclear protein transcription factorFOG1[9].Studies have shown thatFOG1can inhibit the activity ofGATA-1[10]. This may result in lower transcriptional activity ofGATA-1than forGATA-2.Therefore, the combination of theTERTpromoter andGATA-1can reduce the expression of theTERTmRNA. Finally, the mutant A allele of rs2736109 may lead to shorter telomere lengths. The telomere length in the AA genotype was significantly lesser than that in the GG+AG genotype, which isconsistent with the results of this study.
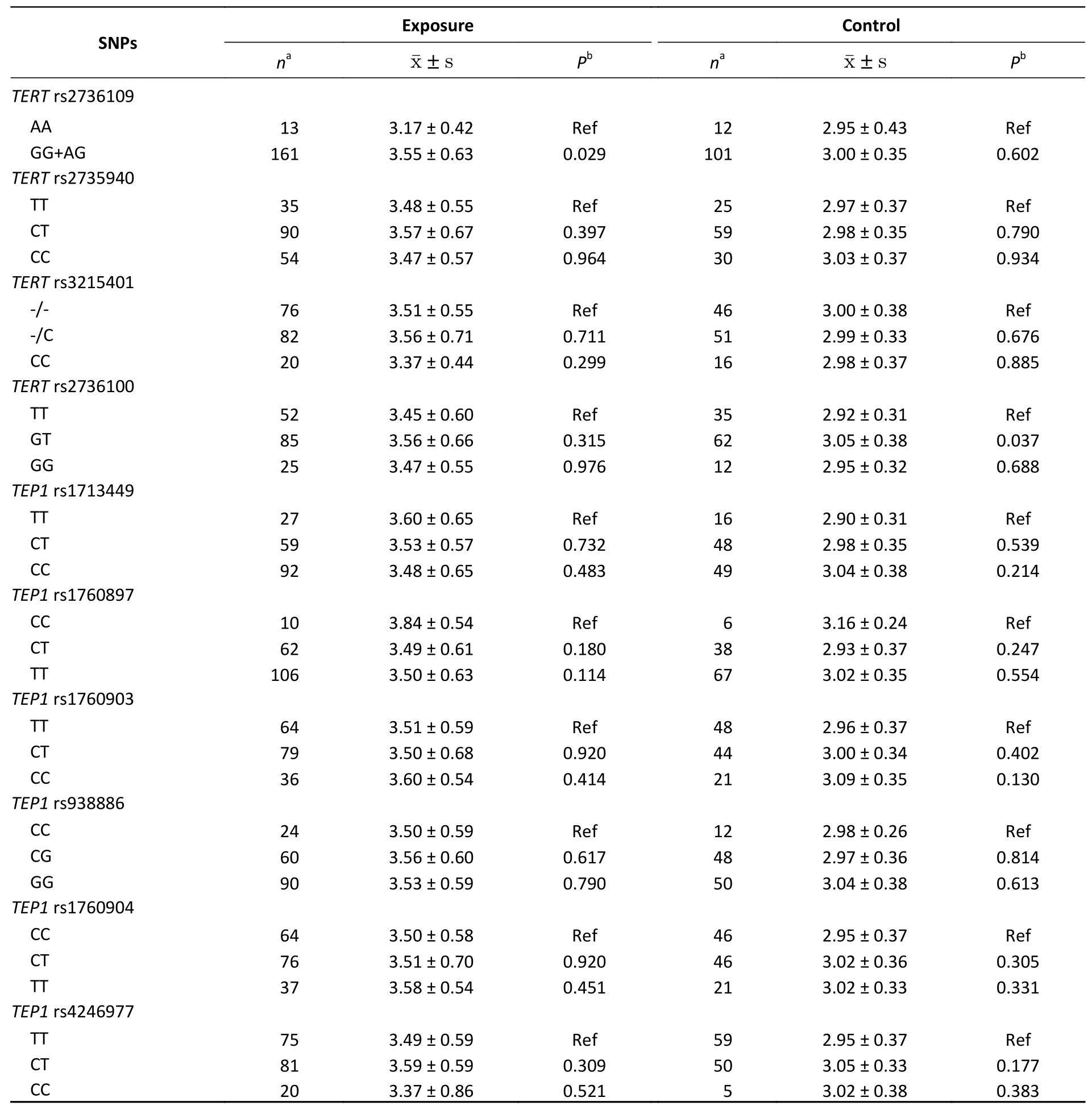
Table 1. Telomere length for polymorphisms in the genes TERT and TEP1

Table 2. Factors influencing telomere length
To our knowledge, this is the first study to investigate the relationship betweenTERTandTEP1polymorphisms and telomere length in workers exposed to omethoate. Our study has several limitations. First, it is a cross-sectional design and therefore does not address the temporal or causal relationship between omethoate exposure and telomere length decrease. Further follow-up is required to confirm these relationships. Second, the molecular mechanisms of the selected SNPs in telomere length shortening remain unclear, and cellbase experiments are required to elucidate these mechanisms. Finally, factors that might affect telomere length, such as chronic diseases and inflammatory conditions, were not considered due to limited information.
In summary, we explored the relationship between telomerase gene (TERTandTEP1)polymorphisms and telomere length in long-term low-level omethoate-exposed workers. TheTERTrs2736109 polymorphism is the main factor affecting telomere length. Through this study, the molecular mechanism of organophosphorus pesticides that cause telomere prolongation was further explored.Our study provides a basis to screen for workers susceptible to occupational exposure. This is conducive to improving workers' health protection and reducing occupational contact damage.
Conflicts of InterestNone.
Author ContributionsCHENG Shuai and LIU Bin wrote the manuscript; YANG Yong Li and GUO Zhi Feng analyzed the data; DUAN Xiao Ran performed the experiments; LIU Su Xiang and LI Lei contributed to specimen collection; YAO Wu and WANG Wei contributed constructs; WANG Wei designed the experiments. All authors commented on the article before submission.
AcknowledgmentsThe authors are grateful to all the individuals who volunteered for the study.
Ethical Conduct of ResearchThe authors state that they have obtained appropriate institutional review board approval or have followed the principles outlined in the Declaration of Helsinki for all human or animal experimental investigations. In addition,for investigations involving human subjects,informed consent was obtained from the participants involved.
&These authors contributed equally to this work.
#Correspondence should be addressed to WANG Wei,Tel: 86-371-67781466, E-mail: ww375@zzu.edu.cn
Biographical notes of the first authors: CHENG Shuai,male, born in 1993, MPH, majoring in occupational cancer and biomarkers; LIU Bin, male, born in 1992, MPH,majoring in occupational cancer and biomarkers.
Received: December 21, 2020;
Accepted: March 22, 2021
 Biomedical and Environmental Sciences2021年10期
Biomedical and Environmental Sciences2021年10期
- Biomedical and Environmental Sciences的其它文章
- Deep Learning in Chest Radiography: Detection of Pneumoconiosis*
- lsolation and Characterization of Vagococcus fluvialis from Bats*
- Cerebrospinal Fluids from Patients with Five Common Genetic Prion Diseases in China Display Distinct Reactivities in the RTQulC Assays*
- Molecular Spectrum of α- and β-Thalassemia among Young lndividuals of Marriageable Age in Guangdong Province,China*
- Obesity rather than Metabolic Syndrome is a Risk Factor for Subclinical Hypothyroidism and Thyroid Autoimmunity*
- The Combined Effect of Dyslipidemia on the lncidence of Type 2 Diabetes: A Prospective Cohort Study in Northwest of China*