
Alpl 基因缺乏抑制正畸牙移动的实验研究
2021-11-05 09:09张立书刘安琪
口腔颌面外科杂志 2021年5期
张立书 , 张 琦 , 刘安琪 , 金 钫
(1. 军事口腔医学国家重点实验室,国家口腔疾病临床医学研究中心,陕西省口腔疾病临床医学研究中心,第四军医大学口腔医院口腔正畸科,陕西 西安 710032; 2.中国人民解放军陆军第71 集团军医院,江苏 徐州 221000)
牙齿受力后, 牙周膜和牙槽骨发生活跃的组织改建是牙齿移动的生理基础[1],从细胞层面观察,压力侧破骨细胞生成增多,吸收压力侧的牙槽骨使牙齿向压力侧移动, 同时张力侧成骨细胞生成增多,形成类骨质,使在张力作用下拉伸变宽的牙周膜恢复到正常牙周膜宽度[2-4],但其具体的分子机制仍未明确。 因此,牙周组织受力后,介导牙齿移动的具体机制仍是目前正畸领域的研究热点。
Alpl 基因参与编码组织非特异性碱性磷酸酶(tissue-nonspecific alkaline phosphatase,TNALP),而TNALP 是碱性磷酸酶(alkaline phosphatase,ALP)的一种,主要在骨和肝脏组织中表达,在牙齿和骨骼系统发育中发挥关键作用[5]。 ALP 在成骨细胞系中表达,可作为成骨分化的重要标志物,在间充质干细胞(mesenchymal stem cells,MSCs)成骨分化的早期,ALP 表达量开始升高, 其表达量的高低可作为判断成骨细胞分化程度和功能的指标[6]。 正畸牙移动中张力侧的骨形成与ALP 活性密切相关,但由于缺乏相应的模式动物,目前仍然缺乏ALP 表达减少是影响正畸牙移动的直接体内证据。 本研究通过构建Alpl 半敲除的转基因小鼠, 首次观察到Alpl 基因半敲除后,小鼠牙齿移动受到抑制,为体内探讨牙齿移动和牙槽骨改建的机制提供了理论依据。
1 材料和方法
1.1 材料
转基因小鼠 B6.129S7-Alpltm1Sor/J (编号:002741),简称Alpl 小鼠,购于美国杰克逊实验室,4%多聚甲醛(赛驰生物科技有限公司,中国),裂解液(Vigen Biotec 公司,美国),山羊血清(博士德生物工程有限公司, 中国),17%乙二胺四乙酸(ethylene diamine tetraacetic acid, EDTA,Pulpdent 公 司 ,美国),直径为0.1 mm 的不锈钢丝(天天齿科器材有限公司,中国),复合树脂(3M 公司,美国),正畸测力计(天天齿科器材有限公司,中国),镍钛螺旋拉簧(北京有研特材科技有限公司,中国),冰冻切片机(Leica 公司, 德国),HE 染色试剂盒(Leica 公司,德国),ALP 抗体 (Bio-Techne 公司,美国;1∶100),羊抗小鼠二抗(Jacson 公司,美国;1∶200),Hoechst 细胞核染料(Med Chem Express 公司,美国),micro-CT扫描及三维重建系统(Siemens 公司,德国),激光共聚焦显微镜(Nikon 公司,日本)。 小鼠基因鉴定引物序 列 如 下 :IMR0137-5′-CCGTGCATCTGCCAGTTTGAGGGGA-3′ -突 变 型 ;IMR0138-5′ -CTGGCACAAAAGAGTTGGTAAGGCAG-3′-野生型;IMR0139-5′-GATCGGAACGTCAATTAACGTCAAT-3′-通用型。
1.2 方法
1.2.1 Alpl 半敲除小鼠的鉴定 小鼠出生后3~4 周,剪去其趾尖,对小鼠进行标号和基因型鉴定。 首先加入裂解液, 于56 ℃水浴过夜后对趾尖组织进行裂解,12 300 r/min 离心 30 min 后取上清液, 提取DNA 并测定浓度,常规聚合酶链式反应(polymerase chain reaction,PCR)扩增目的基因,琼脂糖凝胶电泳检测目的基因的表达情况,由于敲除Alpl 基因的纯合子小鼠具有胚胎致死性,因此,本实验选用半敲除的杂合子小鼠进行后续实验。 常规实时定量聚合酶链式反应 (real-time quantitative polymerase chain reaction,RT-qPCR)检测血清中 Alpl 基因在野生组和Alpl 半敲除组中的表达量差异。 分别取8~10 周野生型小鼠(野生组)和Alpl 半敲除的杂合子小鼠(Alpl 半敲除组)各12 只用于后续实验,所有实验动物均在第四军医大学实验动物中心繁育,每日12 h 光照循环,造模前正常饮食和饮水,造模后每日喂软食,禁咬硬物。 动物实验经第四军医大学动物伦理委员会批准(批准号:SYXK2012-0023)。
1.2.2 构建正畸牙移动模型 将实验动物分为野生组和Alpl 半敲除组2 组, 分别建立正畸牙移动模型,方法参考本课题组既往报道[7]。 具体如下:常规麻醉后将小鼠仰卧固定于手术台上,用自制开口器撑开其上下颌,暴露上颌磨牙,用乙醇棉球充分擦拭小鼠左侧上颌第一磨牙和上颌切牙,将自制加力装置的一端结扎并用树脂粘接固定于左侧上颌第一磨牙, 另一端同样结扎并用树脂固定于上颌切牙,使用正畸测力器测量30 g 拉力。造模后喂软食,每日检查加力装置脱落情况并记录体重,同时观察小鼠饮食和活动情况。 加力第7 天取上颌骨,于4%多聚甲醛中固定。
1.2.3 Micro-CT 检测 应用micro-CT 分别对野生组和Alpl 半敲除组小鼠左侧上颌骨进行扫描和三维重建。 观察并比较2 组上颌第一磨牙近中移动的距离,根据前期文献中的方法[8],测量咬合面三维重建图中上颌第一磨牙远中边缘嵴中点到上颌第二磨牙近中边缘嵴中点之间的距离,每个样本重复测量3 次后取平均值纳入统计。
1.2.4 HE 染色 样本脱矿后用石蜡包埋并切片,常规脱蜡至水后用苏木素染液染色5 min, 蒸馏水洗净染液,氨水处理3 s,1%盐酸乙醇处理1 s 后水洗终止反应,加入伊红染液染色20 s,蒸馏水洗净染液后将切片过梯度乙醇脱水, 二甲苯中透明10 min,中性树脂封片。
1.2.5 免疫荧光检测 样本脱矿完成后用OCT 包埋液包埋样本,冰冻切片机切片。 将切片置于磷酸盐缓冲液(phosphate buffered saline,PBS)中,以多用途摇床洗3 遍,每次5 min,山羊血清封闭30 min,将ALP 抗体按 1∶100 稀释后加入, 置 4 ℃冰箱内孵育过夜, 第2 天取出后室温下复温30 min,PBS 洗净一抗,按1∶200 的比例稀释二抗,避光条件下室温孵育 1.5 h 后用PBS 洗净二抗,Hoechst 染液染核20 min,PBS 洗净后用80%甘油封片。
2 结果
2.1 成功繁育Alpl 半敲除小鼠
对所繁育的小鼠进行基因型鉴定,凝胶电泳结果显示:野生型小鼠(野生组)为1 条位于167 bp 的电泳条带,Alpl 半敲除的杂合子小鼠(Alpl 半敲除组)为双条带,分别位于167 bp 和400 bp 处(图 1A)。琼脂糖凝胶电泳结果与美国杰克逊实验室提供的预测结果一致,提示本研究成功繁育Alpl 半敲除小鼠。 RT-qPCR 检测可见,Alpl 基因在 Alpl 半敲除组中的表达量与野生组相比,显著下降(图1B)。 大体观察发现,Alpl 半敲除组小鼠体型和活动情况与野生组无明显差别,可进行正常繁育,与美国杰克逊实验室官网提供的小鼠品系描述一致(图1C)。
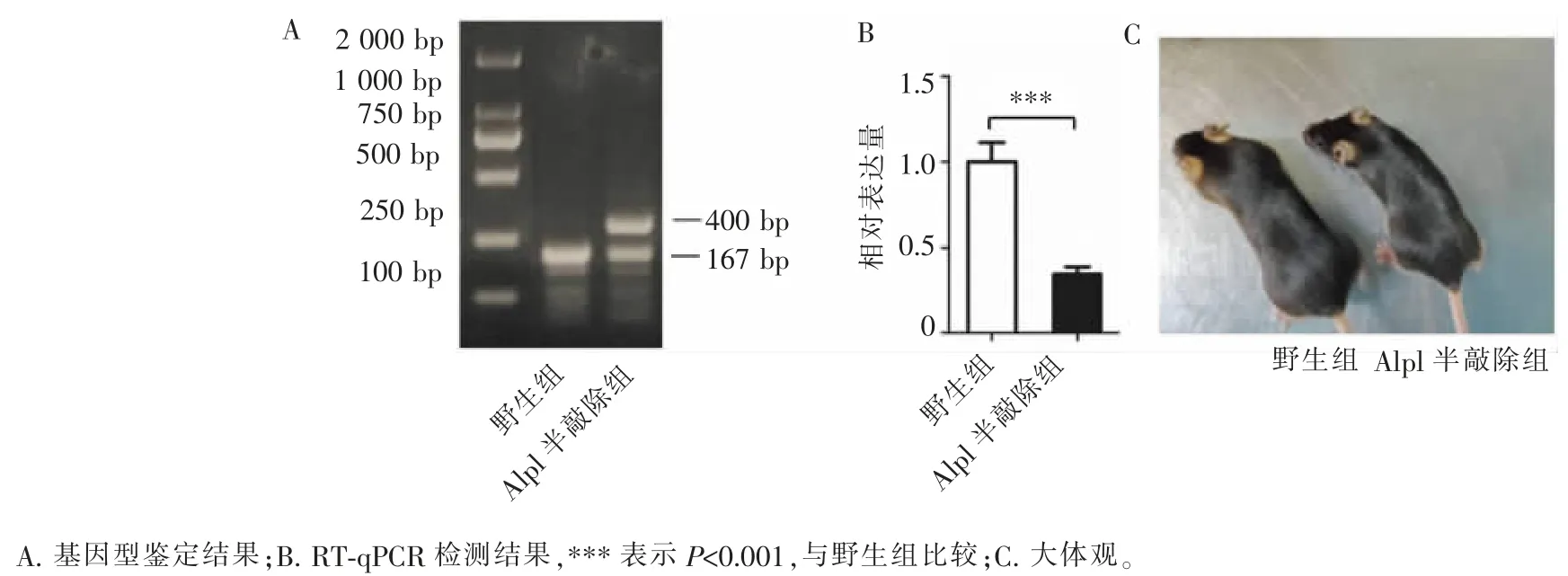
图1 Alpl 小鼠基因型鉴定Figure 1 Genotyping results of Alpl mice
2.2 构建正畸牙移动模型
对野生组和Alpl 半敲除组小鼠上颌第一磨牙加力,每日观察加力装置固位情况,并记录小鼠体质量。 本研究中,2 组小鼠的加力装置均固位良好,无脱落(图2A)。 野生组和Alpl 半敲除组小鼠在正畸加力后进食减少,体质量在加力后前4 天明显下降,而后饮食逐渐恢复,体质量缓慢增加,2 组小鼠体质量变化差异无统计学意义(图2B)。
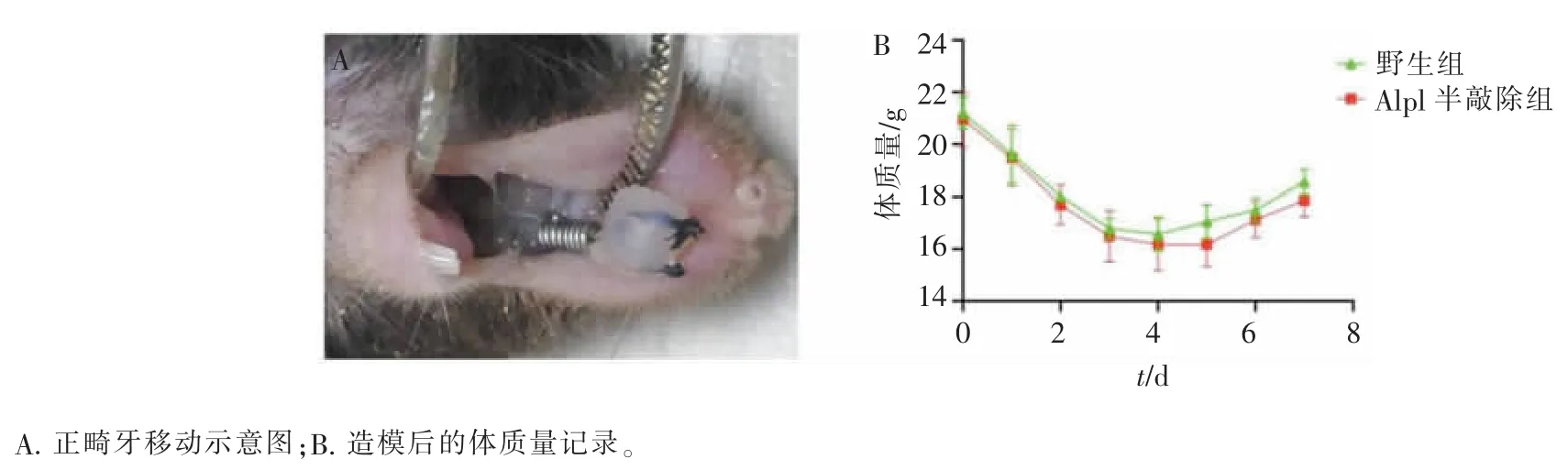
图2 构建正畸牙移动模型Figure 2 Establishment of orthodontic tooth movement model
2.3 Alpl 半敲除能抑制正畸牙移动
Micro-CT 分别扫描并重建野生组和Alpl 半敲除组小鼠上颌骨,观察并比较2 组上颌第一磨牙在加力7 d 后的移动距离。Micro-CT 三维重建结果示,2 组上颌第一磨牙均发生近中移动, 提示本研究中的正畸牙移动模型构建成功(图3A~D)。 此外,通过比较发现,Alpl 半敲除组小鼠上颌第一磨牙近中移动距离和野生组相比明显减少(图3E),提示Alpl基因半敲除能抑制正畸力作用下的牙齿移动,但具体机制仍需进一步探讨。

图3 Alpl 半敲除后,小鼠正畸牙移动受到抑制Figure 3 The orthodontic tooth movement was inhibited after semi-knockout of Alpl
2.4 组织学观察
HE 染色观察野生组和Alpl 半敲除组小鼠上颌第一磨牙的横切面,2 组小鼠牙根压力侧均可见牙周膜受挤压而缩窄,张力侧牙周膜受拉伸而宽度增加。但与野生组相比,Alpl 半敲除组小鼠牙根压力侧未见大量破骨吸收陷窝,且张力侧细胞量较少(图4)。

图4 组织学观察Figure 4 Histological observation
2.5 Alpl 半敲除后,ALP 在正畸牙移动中表达减少
用免疫荧光染色法对Alpl 基因的编码蛋白ALP 进行检测,结果显示,野生组小鼠牙根的张力侧和压力侧均有 ALP 表达(图 5A~C,5G),提示ALP 不仅参与张力侧的成骨分化过程,也在压力侧的骨改建中发挥作用。而Alpl 半敲除组的牙根压力侧和张力侧仅可见ALP 的少量表达(图5D~F,5H),提示Alpl 半敲除后,ALP 在正畸牙移动中表达显著降低, 推测Alpl 基因通过其编码蛋白ALP 影响牙槽骨在正畸力作用下的骨改建,其具体机制尚需进一步研究。
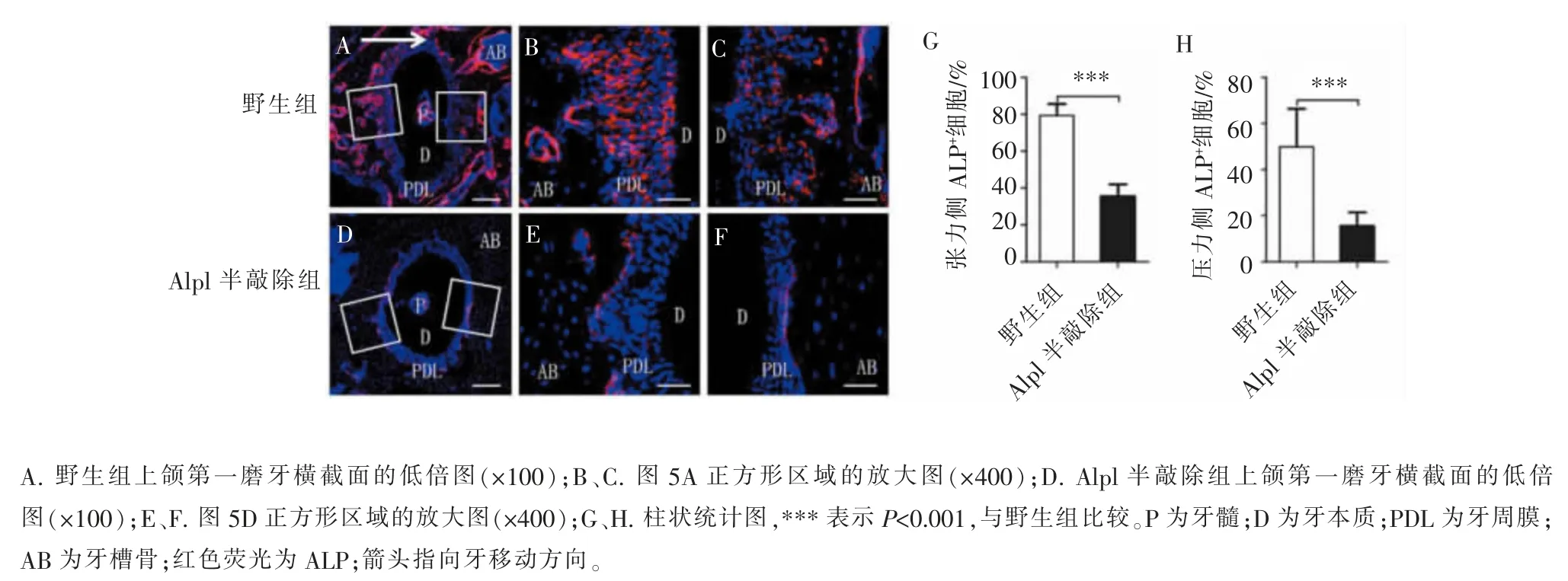
图5 Alpl 半敲除后,ALP 在正畸牙移动中的表达减少Figure 5 ALP expression decreased in the process of orthodontic tooth movement after Alpl semi-knockout
3 讨论
正畸牙移动的过程是牙周组织在正畸力作用下发生改建使牙齿向加力方向移动的过程,同时受免疫系统和循环系统的影响,涉及多种类型细胞行为学改变和复杂分子调控机制, 正畸牙移动的过程是目前正畸领域的研究热点[1,9-10]。 本研究通过使用体内直接半敲除Alpl 基因的小鼠,构建小鼠正畸牙移动模型,对其观察并发现,半敲除Alpl 小鼠在加力7 d 后,正畸牙移动距离减小;同时,Alpl 编码蛋白ALP 在牙周组织中的表达量明显减少,本研究首次通过体内失功能实验明确了Alpl 基因在正畸力作用下,牙槽骨改建过程中的重要作用。
Alpl 基因是ALP 的编码基因,转基因小鼠的发展和应用使人们对Alpl 基因的认识更深了一步[11]。既往研究表明,其编码的ALP 在胚胎发育和骨骼形成中发挥重要作用,Alpl 基因敲除的纯合子小鼠具有围产期致死性,提示ALP 在胚胎发育中的关键作用。 因此,在相关研究中常使用能正常繁育的杂合子小鼠, 这种小鼠体内ALP 合成减少且活性降低,表现为不同程度的骨骼发育缺陷[12],然而,Alpl 基因在正畸领域的研究目前还未见报道。 本研究中所使用的Alpl 半敲除小鼠购自美国杰克逊实验室,该小鼠Alpl 基因启动子后的外显子被β-半乳糖苷酶和新霉素基因所替代,靶向抑制ALP 的合成,常用于人类Alpl 基因突变导致的低碱磷酯酶症(Hypophosphatasia, HPP)的相关细胞和分子机制研究[13-14]。由于敲除Alpl 基因的纯合子小鼠具有胚胎致死性,因此,本实验中Alpl 小鼠均为杂合子。 琼脂糖凝胶电泳结果显示, 杂合子小鼠表现为1 条野生型、1 条突变型的双电泳条带。 RT-qPCR 结果示,Alpl 半敲除组小鼠Alpl 基因表达量明显下降,提示Alpl 杂合子小鼠繁育成功,可进行进一步研究(图 1)。
牙周组织是牙齿移动的生理基础, 临床工作中,我们发现缺乏完整牙周组织的牙齿不能发生移动。 由于正畸牙移动的体内条件复杂,体外无法完全模拟,因此,建立动物正畸牙移动模型对深刻理解体内多因素条件下牙周组织对正畸力的真实反应具有重要意义[3,15]。由于加力装置的粘接固位需要一定的操作空间,因此,口腔操作空间较大的大鼠和比格犬为常见的模式动物, 用于正畸牙移动的研究[16]。随着转基因技术的发展,从细胞和分子水平探讨组织器官的功能及作用模式成为研究的热点。由于转基因技术的限制,转基因小鼠仍是目前主流的动物模型,被广泛用于分子水平的研究。 而小鼠由于口腔内操作空间有限,造模难度大,在正畸领域仅见少量报道[17]。本课题组前期通过实验摸索,成功构建了小鼠正畸牙移动模型,通过物理结扎和化学粘接2 种固位方法,大大提高了加力装置在口内的留存率[7]。Micro-CT 结果(图 3)显示,小鼠上颌第一磨牙向近中移动;HE 染色结果(图4)显示,压力侧牙周膜压缩变窄,张力侧受牵张变宽。 这些结果与既往研究结果一致,提示本实验成功构建小鼠正畸牙移动模型,为进一步观察牙周组织在加力后的改建情况提供了可靠的基础。
牙周组织中的干细胞在响应正畸力和参与牙周改建中发挥了重要作用。 ALP 作为MSCs 向成骨细胞分化的经典标志物, 在成骨细胞系中表达,其表达量的高低可用于评价MSCs 的成骨分化情况[18]。牙周膜干细胞 (periodontal ligament stem cells,PDLSCs)为一群来源于牙周膜的MSCs,体外研究中发现,PDLSCs 在受到张力后,ALP 表达量升高[19]。在实验性动物正畸牙移动模型中,内源性ALP 活性检测常作为牙根张力侧骨形成的检测指标之一[6]。 然而, 目前仍然缺乏Alpl 基因及其编码蛋白ALP 在正畸牙移动中的功能性证据。由于敲除Alpl 基因的纯合子小鼠具有胚胎致死性,因此,本研究中的Alpl小鼠均为杂合子, 尽管未完全消除Alpl 基因的影响,但本研究首次发现半敲除Alpl 基因可抑制正畸加力7 d 后的牙齿移动(图3),同时本研究发现牙周组织中ALP 的表达量与野生型小鼠相比明显下降(图5),推测Alpl 基因通过其编码蛋白ALP 影响正畸力介导的牙周组织和牙槽骨改建。 由于小鼠磨牙较小,且压力侧和张力侧细胞对正畸力的反应并不相同,体外准确提取压力侧或张力侧牙周组织中细胞内的基因和蛋白并进行对比检测十分困难。 因此,后期仍需引入先进的实验方法,进一步设计实验,明确Alpl 基因完全敲除后,牙槽骨改建受到的影响。
综上所述, 本研究通过使用体内直接半敲除Alpl 基因的转基因小鼠, 构建正畸牙移动模型,通过与野生型小鼠对比,发现Alpl 半敲除后,小鼠磨牙的移动距离减少,牙周组织中编码蛋白ALP 表达减少, 推测Alpl 基因在正畸牙移动中发挥重要作用,为进一步探讨正畸牙移动中的细胞和分子机制提供了动物模型和理论依据。
猜你喜欢
口腔医学(2022年10期)2022-11-15
中国药学药品知识仓库(2022年8期)2022-05-09
锦州医科大学报(2022年2期)2022-05-07
口腔医学(2021年10期)2021-12-02
口腔医学(2021年10期)2021-12-02
昆明医科大学学报(2021年5期)2021-07-22
天津医科大学学报(2021年2期)2021-03-29
影像研究与医学应用(2020年24期)2021-01-22
数理医药学杂志(2020年10期)2020-10-17
中国现代医生(2015年8期)2015-04-17
