
盐处理下旱生植物沙芥蛋白激酶相关基因的差异表达分析
2021-10-30 02:44:28汪芳珍杨成行何子华林子茹曾浩源马清
草业学报 2021年10期
汪芳珍,杨成行,何子华,林子茹,曾浩源,马清
(兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业科技学院,甘肃兰州730020)
我国西北荒漠、半荒漠地区气候干旱、降水稀少、蒸发量大,长期以来,人工灌溉是该地区农业生产中主要的补水措施[1]。虽然人工灌溉在短期内可以迅速提高土壤的表层含水量,但由于风沙和高温天气的频发,土壤深层盐分往往会随强烈的水分蒸发转移到地表,造成根际土壤发生次生盐渍化[2-6]。因此,土壤次生盐渍化问题已成为制约我国西北地区农牧业发展和生态恢复的主要因素之一[1]。大多数农作物和牧草长期在耕作条件下栽培种植,其抗盐性的遗传潜力十分有限,这也是土壤盐渍化限制农牧业生产的重要原因。从严酷环境中寻求具有极强抗逆能力的植物,挖掘其抗逆基因资源用于农作物及牧草抗逆性遗传改良已成为目前国际上的研究热点[7-8]。沙芥(Pugionium cornutum)为十字花科(Cruciferae)沙芥属(Pugionium)多浆旱生植物,主要分布于我国西北和内蒙古西南部的荒漠地带,为我国特有种,具有极强的抗旱性,同时对盐渍环境具有极强的适应能力[9-11]。深入研究其耐盐机制,并挖掘其蕴涵的丰富抗逆基因资源,将为农作物和优良牧草抗逆性遗传改良提供重要的理论依据。已有研究发现,盐胁迫下沙芥体内超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)酶活性显著升高,有助于清除氧自由基进而减轻盐胁迫对膜的过氧化[12];5~25 mmol·L-1NaCl 能显著促进沙芥的生长,且随着盐浓度增加,其体内主要渗透调节物质,如Na+、Cl-、SO42-、PO43-等含量均升高,以提高盐胁迫下植株的渗透调节能力[5-6]。最新研究发现,沙芥在盐胁迫下能通过吸收并区域化大量Cl-至地上部作为渗透调节剂来维持细胞内渗透势的平衡,从而有效降低盐胁迫对沙芥幼苗造成的伤害[13]。针对沙芥上述耐盐生理特征,Cui 等[14]采用转录组学手段对盐胁迫下沙芥重要基因的差异表达情况进行了分析,并筛选了一批可能与沙芥耐盐性相关的候选基因,但主要集中于盐胁迫下沙芥体内离子吸收与转运、活性氧清除系统等相关的功能基因,关于盐胁迫下沙芥体内重要调控基因的表达变化目前仍未见报道。
在高盐等逆境胁迫下,植物会接收并转导胁迫信号,启动体内对逆境胁迫的响应机制,进而调控下游相关功能基因对逆境胁迫的响应,其中蛋白激酶是上述信号转导过程中的一类重要的调控蛋白[15-16]。众多研究表明,在逆境胁迫下,植物通过膜受体蛋白激酶感知外界胁迫信号,随后蛋白激酶发生蛋白磷酸化反应,使信号放大并向下传导,进而激活下游的转录因子,以诱导相关抗逆基因的表达,从而增强植物的抗逆性[17]。鉴于此,本研究在已有的盐处理(50 mmol·L-1NaCl)下沙芥转录组数据的基础上[14],分析了盐处理下沙芥根和地上部中重要蛋白激酶编码基因的差异表达情况,以期进一步揭示沙芥耐盐性的相关分子机制,为农作物和优良牧草抗逆性遗传改良提供重要的基因资源。
1 材料与方法
1.1 试验材料培养及转录组数据的获得
沙芥种子采自宁夏石嘴山市沙芥种植园。选取成熟均一的种子先用75%酒精消毒30 s,再用5% NaClO 灭菌12 min,最后用蒸馏水清洗5 次,避光浸泡24 h。置于28 ℃培养箱中发芽,发芽后以石英砂作为培养基质,浇灌Hoagland 营养液培养[14]。随后选取长势良好且一致的4 周龄沙芥幼苗分别进行如下处理:1)对照处理(CK):用正常Hoagland 营养液培养;2)50 mmol·L-1NaCl 处理:含有50 mmol·L-1NaCl 的Hoagland 溶液处理[14]。处理6和24 h 后分别取沙芥的根和地上部材料置于液氮中速冻,之后提取RNA 分别进行转录组测序,进而获得了50 mmol·L-1NaCl 处理6 和24 h 后沙芥根和叶组织的转录组数据[14]。
1.2 蛋白激酶相关基因差异表达分析
利用Blastx 将已获得的沙芥转录组数据[14]中的Unigene 与Swiss-Prot、NR、KEGG、COG 和GO 数据库进行比对得到相应蛋白激酶基因的功能注释。采用每千个碱基的转录每百万映射读取的reads(reads per kilobase of exon model per million mapped read,RPKM)的方法分析不同处理中沙芥根和地上部蛋白激酶相关基因的表达量[14,18-19]。参照Audic 等[20]的方法进行差异表达基因的筛选,并筛选盐处理与对照间的差异表达基因,其中错误发现率(false discovery rate,FDR)≤0.001,且差异倍数在2 倍以上的基因即为差异表达基因(differentially expressed genes,DEGs)。
2 结果与分析
2.1 50 mmol·L-1 NaCl 处理下沙芥根中蛋白激酶相关基因表达分析
50 mmol·L-1NaCl 处理6 h 后,沙芥根中有18 个富亮氨酸重复类受体蛋白激酶(leucine rich repeat receptorlike kinases,LRR-RLKs)、1 个细胞壁连接的类受体激酶(wall associated kinase-like,WAK)、11 个促分裂原活化蛋白激酶级联途径(MAPK/MAPKK/MAPKKK)相关基因上调表达(图1),其中6 个LRR-RLKs(CL2242.Contig2_All、CL797.Contig29_All、CL6717.Contig2_All、CL7990.Contig5_All、Unigene29560_All、CL1136.Contig10_All)、1 个WAK(CL807.Contig2_All)、2 个MAPK(CL1291.Contig1_All、CL2088.Contig2_All)编码基因在对照处理下不表达,而在50 mmol·L-1NaCl 处理6 h 后的沙芥根中表达(表1),表明LRR 类受体蛋白激酶(LRR-RLK)、促分裂素原活化蛋白激酶(mitogen-activated protein kinase,MAPK)可能参与调控沙芥根系对短期盐胁迫的响应。

图1 50 mmol·L-1 NaCl 处理6 和24 h 后沙芥根中蛋白激酶相关DEGsFig.1 Protein kinase-associated DEGs in root of P.cornutum under 50 mmol·L-1 NaCl for 6 and 24 h
此外,50 mmol·L-1NaCl 处理6 h 后,沙芥根中3 个增强抗病性因子(enhanced disease resistance 1,EDR1)、7个酪蛋白激酶(casein kinase,CK)、1 个糖原合成酶激酶(glycose synthase kinase 3,GSK3)和1 个蛋白激酶C(protein kinase C,PKC)编码基因显著上调表达(图1),其中有1 个EDR1(CL4341.Contig2_All)和1 个CK(CL1663.Contig1_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理6 h 后的沙芥根中上调表达(表1);根中有9 个组成型三重反应基因(constitutive triple response 1,CTR1)显著下调表达(图1),其中,2 个CTR1(CL3516.Contig2_All、CL2980.Contig2_All)在对照处理下均有表达,而在50 mmol·L-1NaCl 处理6 h 后的沙芥根中均不表达(表2)。

表1 沙芥根中对照处理下不表达而50 mmol·L-1 NaCl 处理6 h 后表达的蛋白激酶相关基因Table 1 Genes related to protein kinase expressed in root of P.cornutum under 50 mmol·L-1 NaCl for 6 h but not expressed in root under control condition

表2 沙芥根中对照处理下表达而50 mmol·L-1 NaCl 处理下不表达的CTR1 编码基因Table 2 Genes encoding CTR1 expressed in root of P.cornutum under control but not expressed under 50 mmol·L-1 NaCl treatment
50 mmol·L-1NaCl 处理24 h 后,沙芥根中有14 个LRR-RLKs、1 个WAK 编码基因的表达显著上调(图1),其中3 个LRR-RLKs(CL797.Contig15_All、CL2242.Contig1_All、CL217.Contig3_All)编码基因在对照处理下不表达,而在50 mmol·L-1NaCl 处理24 h 后的沙芥根中上调表达(表3);虽然钙依赖性蛋白激酶(calciumdependent protein kinase,CDPK)编码基因下调表达的数目大于上调表达的数目,但有2 个CDPK(CL5176.Contig7_All、Unigene12783_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理24 h 后被显著诱导(表3)。此外,50 mmol·L-1NaCl 处理24 h 后,沙芥根中有4 个CK、1 个GSK3 编码基因的表达被显著诱导,而8 个CTR1编码基因下调表达(图1),其中3 个CTR1(CL2548.Contig11_All、CL2980.Contig4_All、CL3385.Contig7_All)在对照处理下均有表达,而在50 mmol·L-1NaCl 处理24 h 后的沙芥根中不表达(表2)。此外,1 个CK(Unigene2819_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理24 h 后沙芥根中高丰度表达(表3)。

表3 沙芥根中对照处理下不表达而50 mmol·L-1 NaCl 处理24 h 后表达的蛋白激酶相关基因Table 3 Genes related to protein kinase expressed in root of P.cornutum under 50 mmol·L-1 NaCl for 24 h but not expressed in root under control condition
进一步分析表明,50 mmol·L-1NaCl 处理6 和24 h 后沙芥根中有6 个CTR1(CL2548.Contig6_All、CL101.Contig3_All、CL4149.Contig6_All、CL70.Contig 23_All、CL3225.Contig5_All、CL7357.Contig2_All)编 码基因均下调表达,1 个LRR-RLK(Unigene29671_All)编码基因显著上调表达(表4)。

表4 50 mmol·L-1 NaCl 处理6 和24 h 后沙芥根中均上调或下调表达的蛋白激酶相关DEGsTable 4 Protein kinase-associated DEGs that were upregulated or down-regulated in root of P.cornutum under 50 mmol·L-1 NaCl for both 6 and 24 h
2.2 50 mmol·L-1 NaCl 处理下沙芥地上部蛋白激酶相关基因表达分析
50 mmol·L-1NaCl 处理6 h 后,沙芥地上部有13个LRR-RLKs、2 个WAK、8 个CDPK 编码基因的表达显著上调(图2),其中4 个LRR-RLKs(CL8787.Contig2_All、CL8721.Contig3_All、CL7704.Contig2_All、CL4552.Contig2_All)、1 个 WAK (CL807.Contig2_All)、4 个CDPK(CL707.Contig1_All、CL5176.Contig7_All、CL1773.Contig18_All、CL4470.Contig2_All)编码基因在对照处理下不表达,而在50 mmol·L-1NaCl 处理6 h 后的沙芥地上部中表达(表5);虽然MAPK/MAPKK/MAPKKK 中下调表达的数目大于上调表达的数目,但有2 个MAPK(CL10.Contig5_All、Unigene7805_All)、1 个MAPKK(CL3832.Contig4_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理6 h后被显著诱导(表5)。此外,50 mmol·L-1NaCl 处理6 h 后,沙芥地上部有1 个CTR1 编码基因下调表达,6 个EDR1、5 个CK、1 个PKC 编码基因的表达被显著诱导(图2),其中2 个EDR1(CL5872.Contig4_All、CL4341.Contig2_All)、3 个CK(CL1663.Contig1_All、Unigene16644_All、Unigene9975_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理6 h 后被显著诱导(表5)。
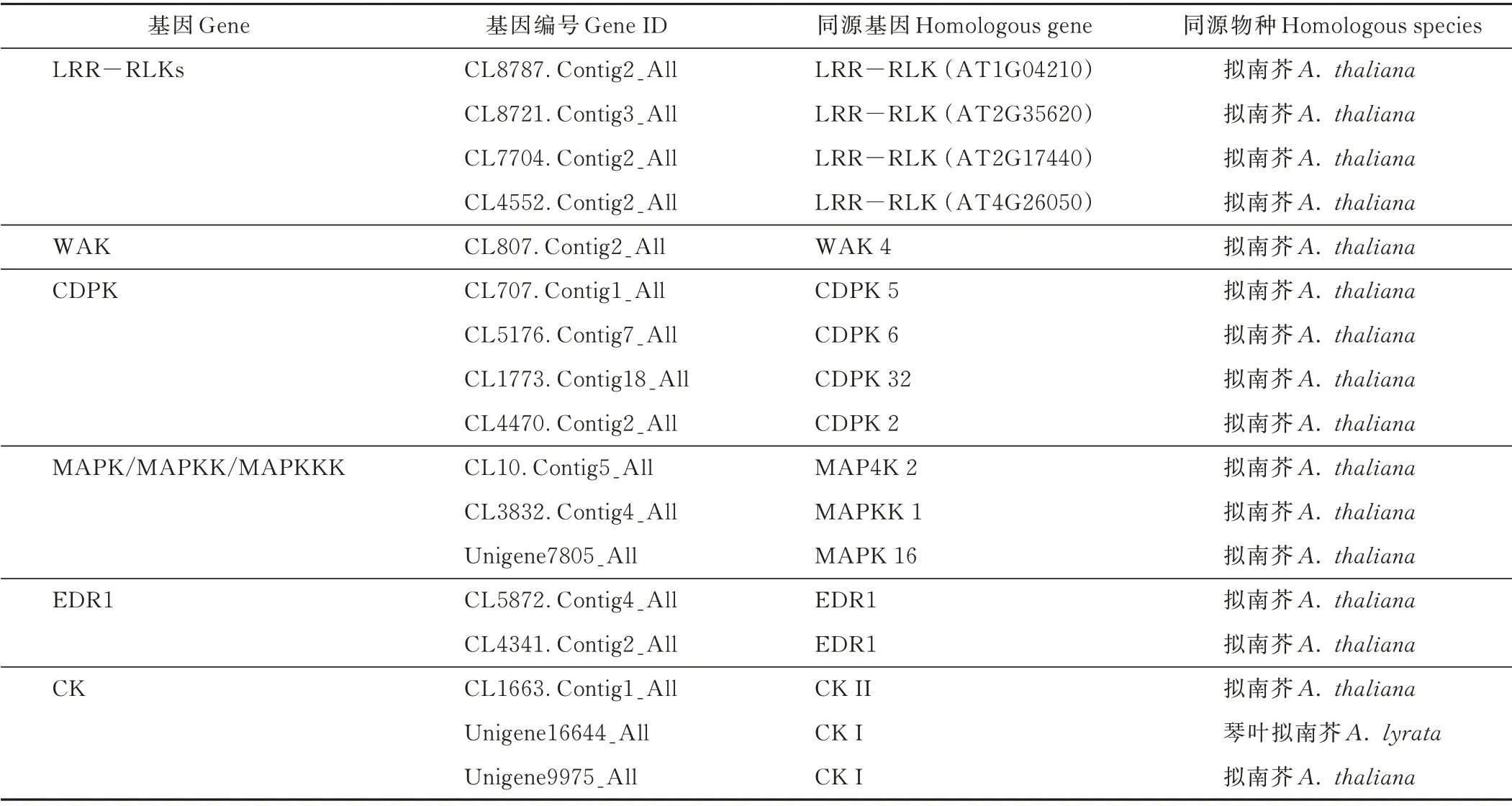
表5 沙芥地上部对照处理下不表达而在50 mmol·L-1 NaCl 处理6 h 后表达的蛋白激酶相关基因Table 5 Genes related to protein kinase expressed in shoot of P.cornutum under 50 mmol·L-1 NaCl for 6 h but not expressed in shoot under control condition

图2 50 mmol·L-1 NaCl 处理6 和24 h 后沙芥地上部蛋白激酶相关DEGsFig.2 Protein kinase-associated DEGs in shoot of P.cornutum under 50 mmol·L-1 NaCl for 6 and 24 h
50 mmol·L-1NaCl 处理24 h 后,沙芥地上部有3 个CTR1 编码基因下调表达,2 个EDR1 编码基因的 表达显著上调(图2),1 个EDR1(CL1578.Contig1_All)编码基因在对照处理下不表达,而在50 mmol·L-1NaCl 处理24 h 后的沙芥地上部中上调表达(表6)。此外,50 mmol·L-1NaCl 处理24 h 后,虽然沙芥地上部LRR-RLK 编码基因中下调表达的数目大于上调表达的数目,CDPK 编码基因中上调表达数目与下调表达数目相同,但有4 个LRR-RLKs(CL3884.Contig5_All、CL4661.Contig4_All、CL3235.Contig3_All、CL4047.Contig1_All)、2 个CDPK(CL707.Contig1_All、CL1773.Contig18_All)在对照处理下不表达,而在50 mmol·L-1NaCl 处理24 h 后被显著诱导(表6)。

表6 沙芥地上部对照处理下不表达而50 mmol·L-1 NaCl 处理24 h 后表达的蛋白激酶相关基因Table 6 Genes related to protein kinase expressed in shoot of P.cornutum under 50 mmol·L-1 NaCl for 24 h but not expressed in shoot under control condition
进一步比较分析发现,50 mmol·L-1NaCl 处理6和24 h 后沙芥地上部有5 个LRR-RLKs(CL797.Contig29_All、CL8787.Contig2_All、CL4552.Contig2_All、CL3884.Contig6_All、CL8721.Contig3_All)、1 个CDPK(CL1773.Contig18_All)、1 个CK(CL5415.Contig8_All)和1 个EDR1(CL1578.Contig8_All)编码基因均上调表达,1 个CTR1(CL3385.Contig7_All)编码基因下调表达(表7)。
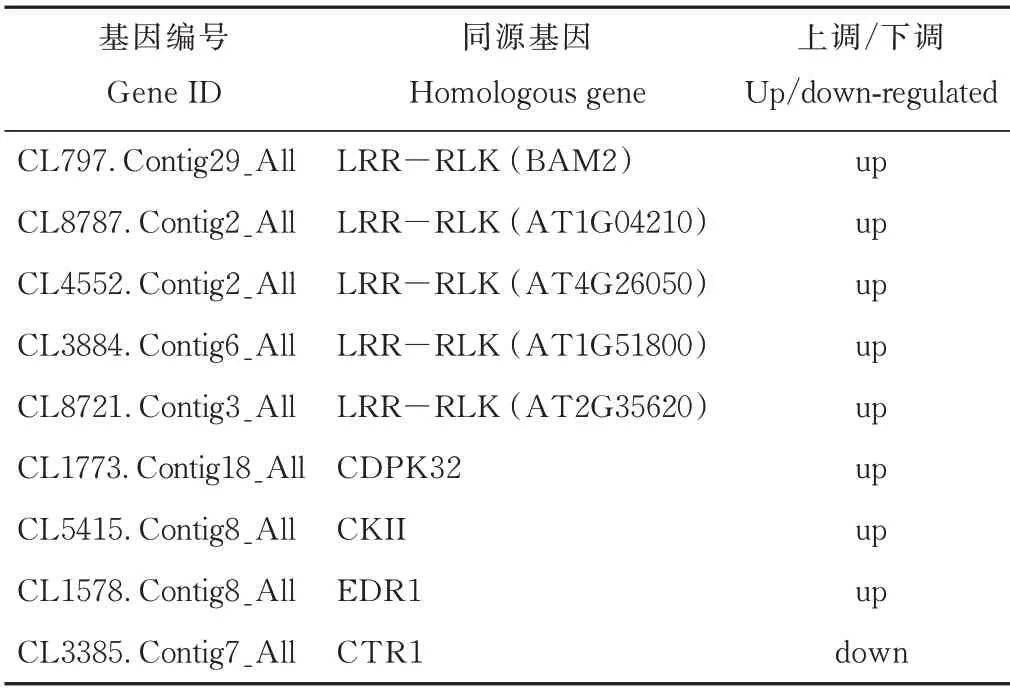
表7 50 mmol·L-1 NaCl 处理6 和24 h 后沙芥地上部中均上调或下调表达的蛋白激酶相关DEGsTable 7 Protein kinase-related DEGs that were up-regulated or down-regulated in shoot of P.cornutum under 50 mmol·L-1 NaCl for both 6 and 24 h
3 讨论
沙芥具有较强的耐盐性,蕴涵着丰富抗逆基因资源[13,21]。已有研究从离子吸收与转运、活性氧清除系统等方面分析了盐处理下沙芥体内的差异表达基因,筛选了一批可能与沙芥耐盐性相关的候选基因[14]。本研究系统分析了盐处理下沙芥根和地上部蛋白激酶相关基因的差异表达,发现50 mmol·L-1NaCl 处理下沙芥根和地上部众多参与调控植物逆境响应的蛋白激酶编码基因的表达发生显著变化,主要包括LRRRLKs、MAPK/MAPKK/MAPKKK 及CTR1 等编码基因。
研究表明,LRR-RLKs 在植物耐盐性中发挥着重要的调控作用[15]。盐胁迫下蒺藜苜蓿(Medicago truncatula)LRR-RLKs 成员Srlk 编码基因快速上调表达,通过调控植株根系的生长发育以调节植物的耐盐性[22]。马铃薯(Solanum tuberosum)StLRPK1通过参与调控水杨酸、茉莉酸、脱落酸等信号通路,从而调控植株对盐胁迫等多种逆境的响应[23]。黄丝瓜藓(Pohlia nutans)PnLRR-RLK基因在拟南芥中的超表达可通过提高AtHKT1、AtSOS3、AtP5CS1和AtADH1等耐盐相关关键基因的表达,从而增强植株的耐盐性[24]。本研究发现,50 mmol·L-1NaCl 处理6 和24 h 后,沙芥根中分别有18 和14 个LRR-RLKs 编码基因的表达显著上调,其中,分别有6 和3 个在沙芥根中对照处理下不表达而在50 mmol·L-1NaCl 处理6 和24 h 后均表达的蛋白激酶相关基因。而在地上部中,50 mmol·L-1NaCl 处理6 和24 h 后,各有13 个LRR-RLK 基因的表达显著上调,其中,各有4 个LRR-RLK 基因在对照处理下沙芥地上部不表达而在50 mmol·L-1NaCl 处理6 和24 h 后表达,且有5 个LRRRLKs 在50 mmol·L-1NaCl 处理6 和24 h 后沙芥地上部中均上调表达。上述LRR-RLKs 可能在沙芥适应盐胁迫过程中发挥着重要作用。
MAPK 家族是丝氨酸/苏氨酸蛋白激酶的一个大家族,家族成员包括MAPK、MAPKK 和MAPKKK 3 种类型,它们通过MAPKKK→MAPKK→MAPK 逐级磷酸化将外来信号级联放大,激活下游相关基因的表达并启动相应的生理生化反应[17]。研究表明,MAPK/MAPKK/MAPKKK 蛋白激酶也参与调控植物对盐胁迫的应答[17]。如研究发现,在盐胁迫下超表达黄瓜(Cucumis sativus)CsNMAPK的转基因烟草(Nicotiana tabacum)种子的萌发率要明显高于野生型[25]。玉米(Zea mays)MAPK 家族基因ZmMKK4超表达能通过增加植株体内脯氨酸和可溶性糖含量、提高POD 和CAT 酶的活性,进而显著提高拟南芥的耐盐性[26]。毛果杨(Populus trichocarpa)MAPK家族成员PtMAPKK4 编码基因的超表达亦可显著增强烟草的耐盐性[27]。在本研究中,50 mmol·L-1NaCl 处理6 h 后沙芥根中共有11 个MAPK/MAPKK/MAPKKK 蛋白激酶基因的表达显著上调,据此可以推测,MAPK/MAPKK/MAPKKK 家族蛋白可能参与调控沙芥对盐胁迫的早期响应过程。
除众多蛋白激酶正向调控植物抗逆性外,还有一些蛋白激酶家族负向调控逆境响应,其中CTR1 是植物耐盐信号传导途径的一类负调控因子[16,28]。在模式植物拟南芥的研究中发现,与野生型相比,盐胁迫下ctr1突变体地上部Na+浓度显著降低而K+浓度显著增加,导致ctr1突变体地上部具有较高的K+/Na+比,从而使得ctr1突变体对盐胁迫的耐受性显著强于野生型植株[29-31]。秘彩莉等[28]将小麦(Triticum aestivum)中的TaCTR1基因在烟草中超表达后,发现转基因植株的耐盐能力明显弱于野生型植株。本研究发现,50 mmol·L-1NaCl 处理下沙芥根中多个CTR1 类蛋白激酶编码基因均下调表达,特别是50 mmol·L-1NaCl 处理6 和24 h 后根中有6 个CTR1 编码基因的表达均显著下调,可以推测,上述CTR1 可能在沙芥根系响应短期盐胁迫过程中发挥着重要的负调控作用。
4 结论
基于已有沙芥转录组数据分析了盐处理下沙芥根和地上部蛋白激酶相关基因的差异表达情况,筛选了可能参与调控沙芥耐盐性的重要蛋白激酶。其中,富亮氨酸重复类受体蛋白激酶(LRR-RLKs)在沙芥适应盐胁迫过程中可能发挥着重要调控作用,促分裂原活化蛋白激酶级联途径(MAPK/MAPKK/MAPKKK)信号通路可能在沙芥响应短期盐胁迫过程中发挥关键作用,乙烯信号转导途径中重要负调控因子CTR1 可能在沙芥根系响应盐胁迫过程中发挥负调控功能。
猜你喜欢
小学生优秀作文(低年级)(2024年4期)2024-05-07 03:13:40
语文周报·教研版(2022年2期)2022-04-27 00:58:07
小学生优秀作文(低年级)(2021年3期)2021-03-15 07:44:26
小学生优秀作文(低年级)(2020年9期)2020-10-26 09:10:34
科学(2020年2期)2020-08-24 07:57:00
河南农业科学(2020年2期)2020-03-11 10:44:04
上海农业学报(2016年2期)2016-10-27 00:49:35
浙江农业学报(2016年7期)2016-06-15 20:30:08
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:17
医学研究杂志(2015年3期)2015-06-10 06:41:52
