
闽楠叶形态与叶脉网络性状关系对城市生长环境的响应
2021-10-29 03:22:50罗丽莹王云龙王光军
生态学报 2021年19期
罗丽莹,陈 楠,王云龙,王光军,*
1 中南林业科技大学生命科学与技术学院,长沙 410004 2 南方林业生态应用技术国家工程实验室,长沙 410004
叶形态性状是指示植物如何适应特定环境的重要指标[1],它能够有效地反映生存环境的变化, 并且影响植物的基本行为和功能[2]。叶脉网络功能性状是叶性状最重要性状之一,是体现叶脉网络结构与叶脉系统功能之间关系结构性状[3],是环境因子、叶脉网络结构和叶脉系统功能三者相互作用的结果表现。叶脉结构具有跨植物物种的共同功能[4],用于机械支持[5]、糖和激素运输[6],补充光合作用过程中因蒸腾而丧失的水分[7],还能通过韧皮部把光合产物和信号分子从叶肉传送到植物其他部分[5]。对于闽楠的形态和功能研究主要集中在生理生态学[8-9]、森林经营[10]、遗传变异[10]、无性繁殖[11]、分布格局[12-13]等,少数人对闽楠叶脉的解剖结构和生态适应性进行了相关研究[14]。目前,大多数有关叶脉网络功能性状相互关系的研究都借助了一定的“媒介”,最常见的是叶片经济性状和叶片水力性状。例如,Sack等[15-16]指出大叶的初级脉直径较大但叶脉密度较小,且叶片水力阻力与中脉轴向阻力正相关,而前者与叶脉密度存在一定关系,后者又取决于木质部的导管数量和直径。John等[17]发现,比叶重与叶脉密度、叶脉直径、单位面积叶脉体积等均有一定的关系。Givnish[18]的研究表明,导管直径变小会降低叶片水分运输效率,但导管数量或叶脉密度可对此进行平衡。
叶脉网络功能性状与叶形态性状是植物适应环境时有较大可塑性的两大性状,它们之间存在着一定的联系。Sack等[15]的研究表明:初级脉直径与叶面积呈正比,初级脉密度与叶面积呈反比,甚至可通过叶片碎块的初级脉密度估算完整叶片面积;而次级脉的直径、密度与叶面积的关系不显著。徐婷等[19]的研究表明:不同坡向的刺槐叶片一级脉密度、二级脉密度均与叶面积呈负相关,并且叶面积的增长速度大于叶脉密度的减小速度。但到目前为止,对上述两类性状关系的研究主要强调叶脉密度与叶面积的关系,而两类性状整体间的关系还鲜有报道。探讨不同生长环境中闽楠叶脉网络功能性状与叶形态性状间的关系,对理解植物为适应环境在形态结构、生理生化方面进行的生态适应策略具有重要意义。
闽楠(Phoebebournei)是国家二级珍稀濒危保护樟科植物[20],其树干端直,冠形优美,枝叶茂密,目前因其独特的生态功能和景观功能,在园林绿化应用中前景广阔[21]。本文对生长于行道和植物园混交林两种不同生长环境的闽楠进行观测,探究叶片4个叶脉网络功能性状与4个形态性状之间的关系,旨在揭示闽楠在城市不同生长环境中的生长特性及其差异,理解闽楠响应异质生境的生态适应机制,为闽楠应用于城市绿化提供理论依据。
1 材料与方法
1.1 样地概况
试验以位于湖南省长沙市中南林业科技大学校园内行道和植物园生长的闽楠(112°59′29″— 112°59′46″E,28°8′7″—28°8′11″N)为研究对象,试验地海拔92—100 m。校区夏天通风,冬季日照充足,排水通畅,属亚热带季风气候,春季多雨,夏秋多晴,严寒期短,暑热期长,年均降水量1361.6 mm,集中于4月,年平均气温为17.4 ℃。本区地层古老,母岩为典型的第四纪网纹红壤,风化程度较深。区内土壤为低酸性土壤或中性土壤,适合植物生长,地带性植被为常绿阔叶林。校区内种植的树种包括常绿及落叶乔木,常绿及落叶灌木,草花植物,水生及藤蔓植物,竹类植物等,近约100科属400余种。
1.2 样品采集与环境因子的测定
2018年8月至2019年7月,试验选取12棵长势良好的闽楠,其中6棵位于主要道路一侧,6棵位于植物园,苗木均于2006年选自湖南永州金洞林场10年生闽楠。初栽时,胸径平均为8.2 cm,树高为6.5 m左右,经12 a的城市环境生长过程,在胸径、树高和外形上均产生明显变异。用高枝剪在每棵树的树冠中上部按东、南、西、北四个方向均匀采样,剪取靠近两年生枝(直径3.5 mm)的成熟无虫害叶片约200 g。天气晴朗时于9:00—11:00,用3415F手持光量子计于4个方位的采样枝条向外5 cm处测定光合有效辐射(PAR),便携式湿度仪测定土壤湿度。行道光合有效辐射为(723.43±67.36) μmol m-2s-1,土壤湿度为0.05 m3/m3,株行距平均为(7.45±0.08) m;植物园混交林光合有效辐射为(584.22±74.24) μmol m-2s-1,土壤湿度为0.12 m3/m3,株行距平均为(4.72±0.32) m。闽楠在两种不同生长环境的生长指标如表1。

表1 两种不同生长环境中闽楠的生长指标
1.3 叶形态和叶脉截面积的测定
每棵树采集的叶片中随机选取叶片30片于Epson Expression 11000XL扫描仪上扫描成图并编号,测量单个叶片的长、宽、面积和周长,测量完形态性状后随机选取8片,沿叶片主脉从上至下分别在叶片的1/3、2/3以及底端处剪取微小矩形块,放入F.A.A固定液中。一段时间后利用LEICA CM1850冷冻切片机切取叶脉纵切面,新制切面放入原固定液浸泡后,选择完好的切片制成装片,用番红-固绿染色后置于显微镜下观察,并用图森IS300显微照相机拍照,之后用ImageJ 1.50i测量主脉面积、主脉韧皮部面积和主脉木质部面积。
1.4 初级脉密度(Major vein density)的鉴定与测量
叶片结构工作组等[22-23]认为,一级脉和二级脉通常被称为初级脉,对叶脉网络起支持和调配作用;三级脉和大于三级的叶脉被称为次级脉,是叶肉细胞与维管束系统进行交换的场所[7,24]。叶脉分级中,一级脉是茎维管束的延续,扩展至整个叶脉长度;二级脉是一级脉的分支[25]。形态性状和叶脉截面积测定完成后,沿叶片主脉从上至下分别在叶片的1/3、2/3处以及底端剪取矩形块(每个矩形块面积为3—5 cm2),扫描测量初级脉的密度。
1.5 数据分析
使用ImageJ 1.50测量叶长、叶宽、叶周长与叶面积,AutoCAD 2012测量初级脉长度,Microsoft Excel 2010对数据进行汇总,并计算相关指标。其中初级脉密度的计算公式为:初级脉密度=初级脉长度/叶片面积;主脉直径、主脉韧皮部直径与主脉直径均为等效直径;可塑性指数=(MAX-MIN)/MAX,MAX和MIN分别代表最大、最小特征值;指标变异系数=标准差/平均值[26]。数据分析前对叶形态、叶脉网络功能性状指标进行单个样本K-S检验,均满足正态分布,使用IBM SPSS Statistics 23.0对数据做描述性统计分析、Pearson相关分析以及典型相关分析,不同绿地环境叶性状平均值的差异比较采用单因素方差分析。运用Sigmaplot 12.5软件中线性函数模型对叶形态组和叶脉网络功能性状组中权重较大的性状进行线性拟合并作图。
2 结果与分析
2.1 闽楠叶形态与叶脉网络功能性状特征
变异系数(Coefficient of variation,CV)和可塑性指数(Plasticity index,PI)是反映进化过程中各性状的保守性或遗传可塑性的两大重要指标,两个指标的值越大,植物对环境的适应性越强[27]。叶形态和叶脉网络功能性状的变异系数统计结果表明(表2),各功能性状的种内变异系数在10.56%—25.44%之间,叶脉网络功能性状变异系数、可塑性指数均表现为植物园环境大于行道环境,而除比叶面积变异系数外,叶形态变异系数、可塑性指数均表现为行道大于植物园。两种生长环境的闽楠叶形态以叶面积的变异幅度最大,叶脉网络功能性状则以初级叶脉密度的变异幅度最大。可塑性指数与变异系数在不同性状间存在差异,两种生长环境的闽楠叶形态性状可塑性指数和变异系数呈现相似规律,由大到小表现为叶面积>叶宽>叶长>叶周长。
将两种生长环境下闽楠叶形态和叶脉网络功能性状特征进行单因素方差分析(表3),表明生长环境对叶形态性状的影响比叶脉网络系统的影响更显著,对叶长、叶宽、叶周长3个叶形态指标特征的影响达到极显著水平(P<0.01),对叶脉网络功能性状指标的影响不显著(P>0.05)。
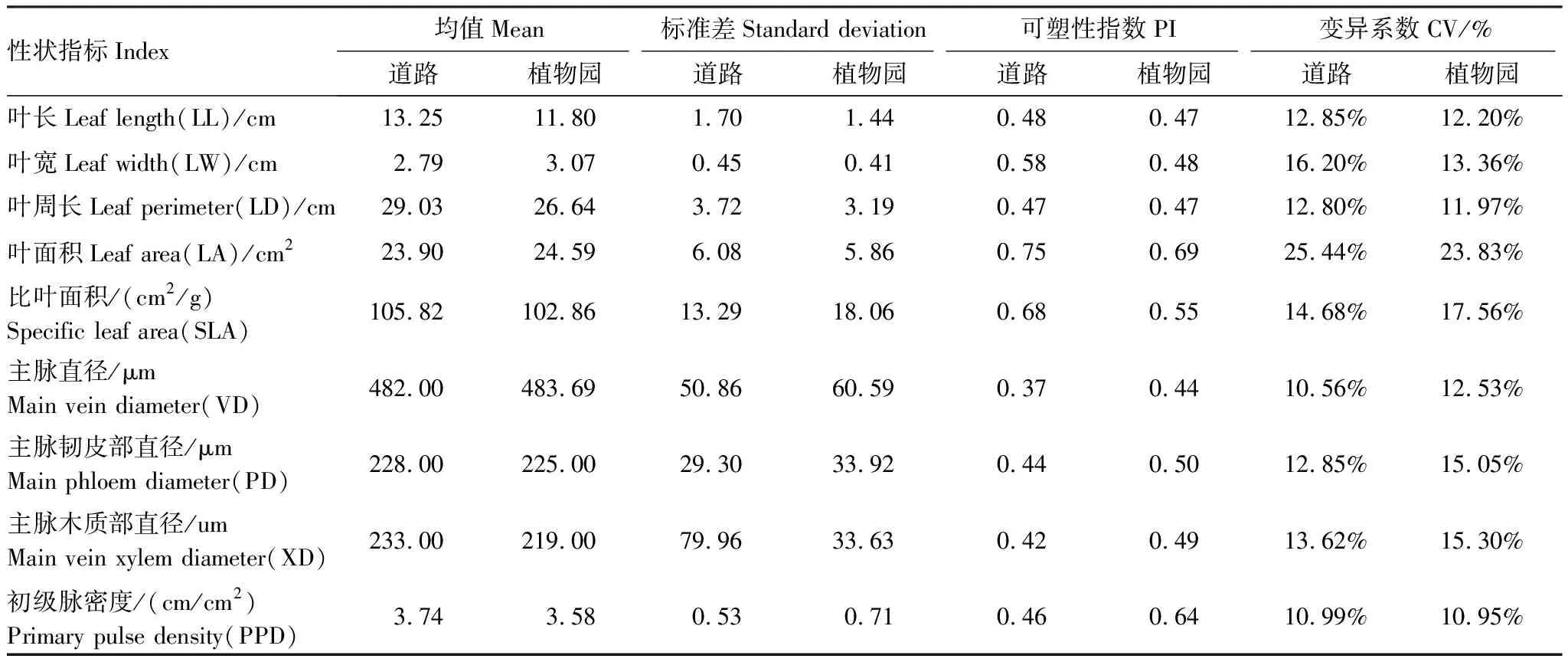
表2 闽楠叶片形态性状与叶脉网络功能性状

表3 两种生长环境叶形态和叶脉网络功能性状的单因素方差分析
2.2 叶形态性状组与叶脉网络功能性状组的典型相关分析
通过将两种生长环境4个叶片形态性状和4个叶脉网络功能性状进行典型相关分析,求得叶片形态和叶脉功能性状两个性状组之间的4个典型相关系数及对应典型变量组合,经Bartlett(χ2)卡方检验,均只有第1典型相关系数达极显著水平,分别为0.804和0.795(图1,图2),说明闽楠形态性状与叶脉网络功能性状之间存在极显著的相关关系。每一个典型变量冗余分析的结果都较大(59.3%,70.3%,77.5%,54.4%),说明各典型变量都能较好地解释相应变量组的变异。
从表4和表5可以看出,对于形态性状组的典型变量U,叶面积(-1.155,-1.269)在两种生长环境中所占权重都是最大,其次是叶长(0.613,0.649),道路闽楠叶片占U1权重最小的是叶周长(0.285),而植物园闽楠叶片占U2权重最小的是叶宽(0.037);对于叶脉网络功能性状组的典型变量V,主脉直径(-0.861,-0.855)在两种绿地环境中占权重均最大,占其权重最小的都是主脉韧皮部直径(-0.053,0.013),说明闽楠形态性状与叶脉网络功能性状这一对典型变量中叶面积与主脉直径的相关最为密切。行道闽楠叶片,与U1相关性最强的是叶宽(λ=-0.958),最弱的是叶周长(λ=-0.601);与V1相关性最强的是主脉直径(λ=-0.979),最弱的是主脉韧皮部直径(λ=-0.687)。对于植物园闽楠叶片,与U2相关性最强的是叶面积(λ=-0.947),最弱的是叶长(λ=-0.808);与V2相关性最强的是初级脉密度(λ=0.882),最弱的是主脉韧皮部直径(λ=-0.624),以上结果表明各性状变量与相应的典型变量之间都具有较强或强烈的相关性。

表4 道路闽楠叶片性状的典型相关分析
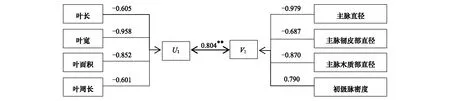
图1 道路闽楠典型变量的典型结构图Fig.1 Canonical structure of the canonical variables of Phoebe bournei around road

表5 植物园闽楠叶片性状的典型相关分析
2.3 不同生长环境叶形态与叶脉网络功能性状的关系
不同生长环境闽楠叶形态性状与叶脉网络功能性状相关性如表6所示,通过Pearson相关分析表明,除植物园闽楠叶片的主脉韧皮部直径与各性状间无显著相关性外,两种环境中的其余性状之间都具有显著相关性。两种生长环境中叶形态性状之间均呈极显著正相关(P<0.01),初级脉密度与叶形态性状、主脉直径呈极显著负相关,表明初级脉密度与叶形态性状、主脉直径呈显著权衡关系。
根据叶形态性状组和叶脉网络功能形状组典型相关分析的结果,选取各性状组权重较大的两个指标进行函数模型拟合分析(图3),分别为叶长、叶面积和主脉直径、初级脉密度,叶长、叶面积与主脉直径存在极显著的正相关关系,其回归方程斜率均表现为植物园闽楠大于行道闽楠,表明叶长、叶面积等叶形态性状与主脉直径存在协同进化关系,随着叶长、叶面积的增加,植物园闽楠叶主脉直径生长速率更明显。叶长、叶面积、主脉直径与初级脉密度呈显著权衡关系,叶形态性状、主脉直径越小叶脉密度越大,且这种权衡关系在两种生长环境中存在差异。
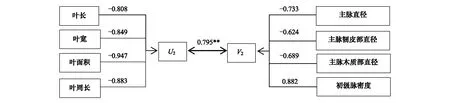
图2 植物园闽楠典型变量的典型结构图Fig.2 Canonical structure of the canonical variables of Phoebe bournei in botanical garden
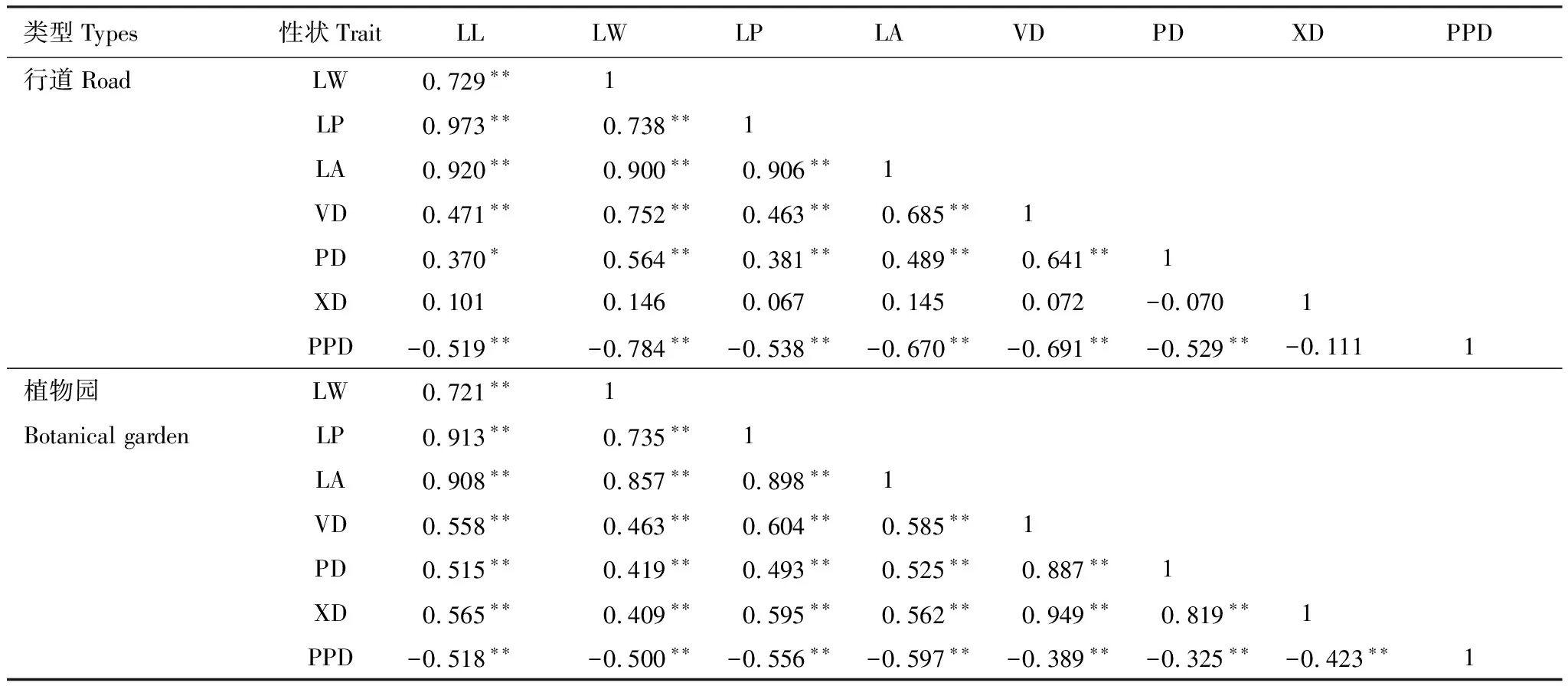
表6 不同生长环境下闽楠叶形态性状与叶脉性状的Pearson相关分析
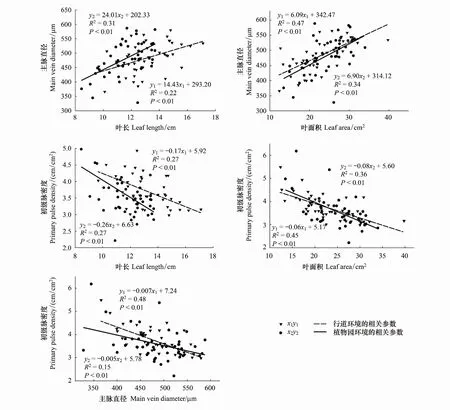
图3 不同生长环境下叶宽、叶面积与叶脉直径、初级脉密度的关系Fig.3 Relationship between leaf width, leaf area, vein diameter and primary vein density in different growing environments
3 讨论
3.1 两种生长环境闽楠叶性状特征及变异程度
叶片作为植物生理活动的重要器官,对异质环境敏感,适应环境过程中表现出一定的表型性状[28]。Philip等[29]的研究认为叶长、叶宽和叶面积均会随着荫蔽程度的加强而增加,本次研究中除叶长的变化情况有差异外都与其相符。竞争很大程度上塑造植物生存环境条件,使其结构和功能不断适应多变的环境,特别是对于幼龄林时期的乔木来说,在响应邻里竞争的过程中,其结构会被强烈塑造[30-31]。王君[32]进行不同水分处理的植物,叶面积和比叶面积会产生显著差异,随胁迫的加深叶面积变小。两种生长环境中,植物园树木密度大,竞争强烈,而行道生长的闽楠株距大,能获得更多的光照,因此生长在植物园环境中更注重叶宽的增加来显著增加叶面积。植物生长环境的差异导致获得性资源发生改变,迫使植株个体在高度、叶片性状、叶脉性状等方面发生可塑性调整[33]。植物园较行道土壤含水率较高,树木密度大遮荫严重,植物园闽楠在土壤水分充足的条件下为了竞争光资源产生最大的光合效益,闽楠选择较低初级叶脉密度、较高叶面积和叶脉直径的叶形态和叶脉网络构建模式来保证植物叶片具有较高的导水率和机械支撑能力。行道土壤含水率较低,且生长于固定面积的树池中,其水资源获取受限,行道闽楠有较大的初级脉密度,以获得更多水分传输路径;闽楠植株之间间距大,枝叶间的遮荫现象小于植物园,光照更充足,为减少蒸腾作用的失水行道闽楠选择较小的叶面积。叶脉网络系统的发育和构型是叶脉结构性状根据生存的环境特点进行经济权衡的结果[34],植物叶形态和叶脉网络功能性状在直接响应环境变化的同时,结合自身生长需求权衡资源获取过程中的投入-收益,对差异化环境产生独特的响应机制,形成最终的叶脉网络功能系统[35],这在一定程度上削弱了环境对叶脉网络功能性状的直接塑造。
Hallik等[36]的研究表明变异系数可反映物种潜在适应能力,功能性状的种间变异系数可高达95%,而种内变异系数则不超过30%。本研究中闽楠各功能性状的种内变异系数在10.56%—25.44%之间,说明各功能性状的变异较为丰富(表2),种内变异虽然较小却是植物对不同生境适应性变化的直接体现。可塑性指数可反映物种克服环境异质性的能力[37],两种环境下叶面积的变异系数和可塑性指数均较大,反映出叶面积在两种环境中都具有极强的可塑性水平,叶形态性状和叶脉网络功能性状受生境异质性的影响,在两种环境中协同变化产生表型差异。
3.2 两种生长环境下叶脉直径与叶脉密度的权衡关系
叶脉具有生理学和力学的双重意义,在生理学上叶脉作为维管束器官提供叶片水分和养分的供应是其主要作用,在力学上作为木质部成分对叶片起支撑作用[3]。在本研究中,主脉直径、主脉韧皮部直径与主脉木质部直径三者之间具有正向的相关性,这是因为叶脉本身就主要由韧皮部与木质部组成,四周是维管束鞘,它们又被包裹在薄壁细胞或厚壁组织中[38],筛管能将光合产物从叶片输送到植物的其他部分,导管能够从叶柄向叶肉输送水分。Mcknow等[39]指出,较大的叶片往往有较大的叶柄和主脉,其中含有更多更大的木质部导管和韧皮部筛管。韧皮部筛管数量多或直径大有助于提高韧皮部运输效率,木质部水力导度与木质部导管数量也与呈正相关关系[38,40],因此韧皮部直径与木质部直径能反应叶片对于光合产物和水分的运输能力。
本研究中初级脉密度与主脉直径具有负相关性,呈显著的权衡关系,这与Sack等[15]对于较大叶片相关研究以及Givnish等[18]的研究结论相同。在植物园混交林中闽楠与其他乔木光竞争,遮阴会降低植物园的土壤水分潜在蒸散且其他树种树干截流雨水改变小气候,使其以较低的叶脉密度、较大的叶脉直径来满足蒸腾作用的水分要求;行道闽楠能获得更多的光但土壤水分含量较低且蒸腾作用强于植物园,为了较大限度利用行道土壤水分并实现光合作用的物质的转化和迁移,就需要较大的叶脉密度来权衡。为了降低与相邻植物的资源和生存空间等的竞争, 植物叶片叶脉密度与叶脉直径存在权衡与协变关系, 改变异质生境中植物形态和生理性状间的相互耦合, 最大限度地达到较高的水分利用效率和光合能力, 以及实现光合作用中的物质迁移与能量转化的生理生化机制[41-42]。
3.3 两种绿地环境下叶形态与叶脉网络功能性状关联性分析
环境因子并非简单地调控某一性状,而是控制植物性状之间的相互关系,通过多个性状的表达来促进功能策略的优化,使性状之间协同地响应环境变化[43-44]。Wright等[45]的研究也指出,气候对某一种叶片性状的影响较小而对不同性状间关系的调控影响较大,并且植物叶片性状之间的关系比植物叶片性状和环境的关系更加紧密。根据特定的环境条件而进化出的形态和生理特性可能是植物在不同生境中生存和繁殖的原因[46-47]。在光合作用和水分运输的权衡下,植物园混交林闽楠为了与其他乔木光竞争,通过减小闽楠的叶脉密度而增加叶片大小的投资来保证光合收益;而行道闽楠则通过更少的叶片资源投入来获取更大的叶脉密度,保证光合所需要的水分,从而叶形态形状组与叶脉网络功能性状组之间协同进化,但不同环境中叶片形态性状组、叶脉网络功能性状组的典型变量与相应原始变量之间的关系表现出差异。为了顺利完成自身的生命过程且实现光合产物投资-收益的最大化,植物会主动调整、转换和补偿叶脉性状的功能及状态[48-49],将生物量优先分配给具有最易获取能力的性状,这是植物提高资源利用效率的生活史对策[4]。植物的生态策略和适应性、响应性功能要通过一系列紧密联系的性状来实现,但性状的构建,稳固和性状之间的强弱变化与替补转换,则靠植物自身的资源权衡机制来调控[50]。例如,不同环境中叶片形态性状和结构功能性状是相互联系的,但其生态策略的表现是一致的[51]。在本研究中,两种不同生长环境下闽楠叶片形态性状组与叶脉网络功能性状组达极显著相关性,对于不同环境的同一种典型变量来说,相应原始变量的权重系数与相关性系数具有较大差异,因此将闽楠作为城市绿化树种时,可以根据绿地环境条件,将叶片形态性状组与叶脉网络功能性状组的相关性作为选种依据,以提高闽楠在城市绿化中的适应性。
4 结论
本文基于城市不同生长环境的闽楠生长,分析闽楠响应异质生境所表现的植物叶形态与叶脉网络功能性状的协同变异与权衡关系,主要得到以下结论:(1)生长环境对叶形态性状的影响比叶脉网络系统的影响更显著,各功能性状的种内变异系数在10.56%—25.44%之间,两种生长环境的闽楠叶形态以叶面积的变异幅度最大,变异系数分别为25.44%和23.83%,叶脉网络功能性状变异系数、可塑性指数均表现为植物园环境大于行道环境,而除比叶面积变异系数外,叶形态变异系数、可塑性指数均表现为行道大于植物园。(2)闽楠叶形态与叶脉性状间存在一定的协调性,并通过调整叶片的结构特征来适应环境的变化。为满足植株获取光合产物和提高水分利用效率的需求,行道闽楠选择了较大的叶脉密度和较小的叶面积来提高水分利用效率;而植物园混交林的闽楠则选择较大的叶面积、较小的叶脉密度和较大的主脉直径,弥补因相邻植物干扰的光竞争,较好的验证了植物对生境异质性的适应机制。(3)闽楠形态性状组与叶脉网络功能性状组之间存在极显著的相关关系。叶形态性状与叶脉直径呈显著正相关,形态性状、叶脉直径与初级脉密度呈显著负相关,行道环境中闽楠以较大的初级脉密度和较小叶面积来确保水分获取和光合之间的平衡;植物园闽楠则采用较低初级叶脉密度、较高叶面积和叶脉直径的叶形态和叶脉网络构建模式,来保证植物叶片具有较高的导水率和机械支撑能力。今后将闽楠应用于城市绿化,还需研究各异质环境影响叶形态和叶脉网络功能构建机制的关键环境因子,根据种植环境条件,将叶片形态性状组与叶脉网络功能性状组的协同变化和权衡关系作为选种依据,以提高闽楠在城市环境中的成活率和适应性。
猜你喜欢
动漫界·幼教365(大班)(2021年11期)2021-11-20 07:49:02
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
南方农业学报(2021年2期)2021-06-30 01:56:03
发明与创新(2020年26期)2020-06-27 07:25:06
湖南文理学院学报(自然科学版)(2020年2期)2020-05-08 08:00:14
新疆农业科学(2020年1期)2020-02-14 03:22:46
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:16
新课程·小学(2019年7期)2019-09-17 12:58:34
江苏农业科学(2019年21期)2019-01-03 02:05:58
红领巾·萌芽(2017年2期)2017-03-09 15:25:16
