
甲壳素及其衍生物在水产饲料中应用
2021-10-23 07:55:52许乐欣王晓清王子奥王红波
湖南饲料 2021年5期
许乐欣 王晓清* 王子奥 王红波
(1.湖南农业大学动物科学技术学院,湖南长沙410128;2.湖南丰尔渔生物科技有限公司,410001)
随着我国经济的快速发展, 人们生活质量日益提高, 水产养殖行业规模的逐渐扩大, 加工行业的技术也日渐成熟, 生产过程中虾类的副产物虾壳量也日渐增多。 大量的虾壳被低价值利用或者直接丢弃, 造成了资源浪费、 环境污染以及经济损失。 而虾壳中有多种可利用的成分, 例如甲壳素, 可以通过提取加工, 应用于水产动物饲料中。
1 甲壳素及其衍生物
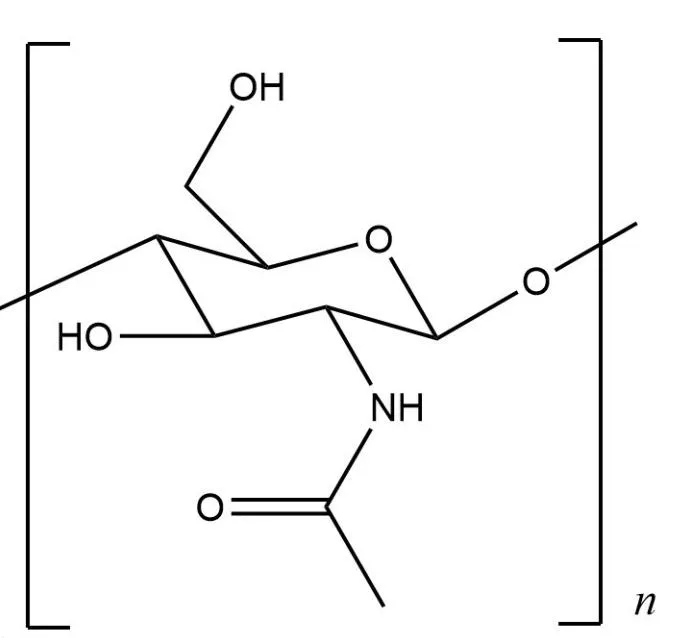
甲壳素(Chitin) 又称为甲壳质、 几丁质,是一种广泛存在于甲壳类动物的多糖物质, 结构如图一所示, 是世界第二大丰富的生物聚合物,仅次于植物纤维。 甲壳素广泛应用于医药行业、工业、 农业。 甲壳素的衍生物有壳聚糖、 壳寡糖、 甲壳胺、 壳聚糖季铵盐等。
图一
壳聚糖(Chitosan) 是甲壳素脱N-乙酰基的产物, N-乙酰基脱去55%以上的就可称之为壳聚糖。 壳寡糖(Chitosan oligosaccharide) 是首先由甲壳素脱乙酰化生成分子质量为几十万到几百万几道尔顿、 吸收率为1%-5%的壳聚糖, 再经生物酶解后得到的分子质量﹤2000u、 可被吸收进入体内的低聚糖, 它由2-10 个氨基葡萄糖通过β-1, 4 糖苷键连接而成, Benchamas 等在研究中发现其具有显著抗菌抗癌等生物活性。 壳聚糖季铵盐是壳聚糖经化学改性修饰得到的衍生物, Li等在实验中发现其可作为疫苗中的佐剂增强免疫原性。
甲壳素及其衍生物的提取工艺也在往高效率低成本发展。 Zhang 等用巴氏醋杆菌和枯草芽孢杆菌从虾壳中提取甲壳素, 工艺较为简单, 操作时间较短, 生产效率高。 Taser 等用类芽孢杆菌从虾壳中提取甲壳素后, 再用氢氧化钠法将提取的甲壳素转化为壳聚糖。 用这个方法得到的壳聚糖分子量较低。 低分子量的壳聚糖具有更高的的溶解度、 更低的毒性。 除此之外, 低分子量的壳聚糖对退行性疾病有抵抗作用。
甲壳素及其衍生物作为饲料添加剂在水产动物上的应用较多,具有调控营养物质代谢、 促进生长、 提高免疫能力等作用,可部分替代传统化学药物及抗生素等使用,推动水产养殖可持续、健康发展。
2 甲壳素的生理功能

甲壳素是阳性趋向剂, 可以促使吸引单核细胞在局部聚集增生演变为巨噬细胞。 另外, 杨怀宇等研究表明甲壳素、 壳聚糖可以活化T 淋巴细胞, 促使T 淋巴细胞释放出各种淋巴因子, 发挥杀死体内微生物的作用, 激活机体的免疫系统。同时甲壳素及其衍生物还可以增强溶菌酶等水解酶活性, 调节机体的免疫功能。

甲壳素及其衍生物具有广谱的抗菌性, 其抗菌抑菌的作用机制对革兰氏阴性菌和革兰氏阳性菌不同, 而且抗菌抑菌的功效因其浓度、 分子量及其脱乙酰程度还有环境的pH 值有所不同。 周锐丽等实验发现甲壳素、 壳聚糖可与微生物的细胞膜作用改变细胞膜的通透性, 从而影响微生物正常的活动, 起到杀菌的作用。 此外, 甲壳素还可与水结合抑制部分酶的活性。
目前的研究表明, 甲壳素及其衍生物具有显著的降低血脂、 调节脂肪代谢的作用。 甲壳素、壳聚糖结构中的葡聚糖胺链含有4 价的铵离子,具有较强的阴离子交换能力, 能够结合带负电荷的胆汁酸, 促使体内胆固醇转化为胆酸, 维持胆酸的储备, 从而降低胆固醇的浓度。
甲壳素、 壳聚糖促进动物生长的原理是其能够改变肠道结构、 提高消化酶的活性, 从而提高机体对营养物质的消化吸收能力。 Chang 等研究表明壳寡糖可以提高黄鱼肉鸡的生长性能。 另外, 甲壳动物可以吸收利用甲壳素及其衍生物,促进本身几丁质酶的分泌, 从而促成蜕壳生长。
3 甲壳素及其衍生物在水产动物饲料中的应用
酸性磷酸酶(ACP) 和碱性磷酸酶(AKP)是生物体内2 种重要的水解酶,能催化磷酸单酯水解, 直接参与体内磷酸基团的转移和代谢,是吞噬细胞杀菌的物质基础,也是甲壳动物机体防御能力的重要组成部分。 丙二醛(MDA) 使机体酶系统和非酶系统产生氧自由基, 导致生物膜中的多不饱和脂肪酸被攻击氧化而形成的脂质过氧化物MDA 浓度的高低间接反映了机体细胞受自由基攻击的严重程度。 超氧化物歧化酶(SOD)是一种抗氧化酶, 能清楚体内过多的超氧自由基。 溶菌酶(LZM) 一种碱性蛋白酶, 广泛存在于鱼类皮肤黏液、 血清、 组织器官及吞噬细胞中, 在鱼类的非特异性免疫防御系统中占重要地位。 抗体是用来判断鱼类特异性免疫能力的主要指标。

目前, 在多种鱼类上的研究表明, 甲壳素及其衍生物在饲料中的添加可以促进鱼类生长, 提高血清内多种酶的活性, 增强免疫力, 提高存活率, 并且降低饲料系数。
李丽鹃等在基础饲料中分别添加0、 25、50、 100mg/kg 的壳聚糖, 投喂平均体重约为7.5g的黄颡鱼42d。 结果显示, 在黄颡鱼的饲料中添加25mg/kg 的壳聚糖显著地提高了黄颡鱼血清中ACP、 AKP、 SOD、 LZM 活性。 肖艳翼等在基础饲料中分别添加0、 1.25、 2.5、 5、 7.5、 10g/kg的壳聚糖, 投喂12g 左右的俄罗斯鲟50d, 测定结果表明一定量的壳聚糖可以提高俄罗斯鲟血清的ACP、 AKP、 SOD、 LZM 活性。 李赫等在基础饲料别添加0、 2.5、 5、 7.5、 10g/kg 的壳聚糖投喂均重为108g 的草鱼, 研究表明5g/kg 的壳聚糖可以增强草鱼血清中AKP、 SOD、 LZM 活性, 并提高补体C3 含量和血液白细胞吞噬活性, 降低MDA 含量。 王荣月等在基础饲料中分别添加0、0.2%的壳寡糖、 0.2%的壳聚糖以及0.2%、 0.1%、0.02%的壳聚糖纳米颗粒, 投喂体质量为(9.910±0.306) g 的红鳍东方鲀幼鱼50d, 研究表明饲料中添加壳聚糖纳米颗粒的效果与壳寡糖的作用效果相当, 可以显著增加肝脏AKP 以及脾脏ACP、 LZM 和肾脏SOD、 LZM 活性, 对鱼体免疫力的提高有一定的促进作用。 蒋锦坤等在基础饲料中添加0、 0.25%、 0.50%、 1.00%和2.00%的壳聚糖投喂虹鳟幼鱼50d, 结果表明,在饲料中添加壳聚糖可以显著提高虹鳟血清中ACP、 AKP、 LZM 活性, 增强虹鳟非特异性免疫能力。 Askarian 等发现在饲料中添加甲壳素, 可以调节大西洋鲑鱼的肠道微生物菌群。 Harikrishnan 等研究发现在饲料中添加甲壳素和壳聚糖有助于提高褐带石斑鱼血液红细胞及白蛋白、 球蛋白含量和吞噬病原体活性, 增强血液免疫能力。 Kumar 等发现饲料中添加甲壳素, 可作为免疫增强剂, 提高露斯塔野鲮对鳃单殖吸虫的抵抗力。 以上研究表明适量的壳聚糖可以作为免疫增强剂可以增强鱼类自身免疫力, 促进鱼类抗体的生成, 提高其抗感染抗病能力。 Kumaran 等用以含有纳米壳聚糖(CSNPs) 的N-乙酰-D-葡萄糖胺 (NAG) 作为添加剂在饲料中分别添加0、0.25、 0.5、 1.0、 2.0g/kg 投喂体质量为 (15.30±0.23g) 的尼罗罗非鱼45d。 实验结果表明1.0g/kg的CSNPs 可以提高尼罗罗非鱼的非特异性免疫。
在促进鱼体生长方面, 主要是能够改变鱼类肠道组织和菌群的情况促进鱼类进食, 以及能够降低鱼的脂肪含量, 增加鱼的可食用率。 王红权等发现在草鱼饲料中添加0.50%壳聚糖能显著促进草鱼生长, 增加草鱼背肌粗蛋白含量, 减少粗脂肪含量, 降低草鱼的投饵系数。 华雪铭等研究发现壳聚糖在饲料中的添加,能够显著降低肌肉脂肪含量,显著提高肌肉蛋白质含量,也能在一定程度上降低肝体比。 0.2%壳聚糖能使肠道淀粉酶活性显著增强(P﹤0.05), 白肌RNA/DNA 显著增大(P﹤0.05), 说明肠道淀粉酶活性增强和蛋白质合成增加是壳聚糖和益生菌促进暗纹东方鲀生长的重要原因。 陆清儿等在三角鲂养殖试验的结果表明添加壳聚糖及其复合物有较好的促长效果,三角鲂平均体增重极显著提高、 饲料系数极显著下降、 成活率明显提高。 孙飞等在含正常豆油、 氧化豆油的基础饲料中分别添加0、 0.02%、0.04%、 0.06%的壳寡糖, 共配制成8 种试验饲料。 结果显示, 在含4%豆油的常规饲料中添加0.02%的壳聚糖可以促进异育银鲫的生长, 改善肠道结构;氧化豆油会对异育鲫鱼的生长和健康造成负面影响, 在该饲料中添加0.04%的壳聚糖可改善由氧化豆油引起的负面影响, 使异育银鲫的健康程度达到正常水平。 Abd El-Naby 等发现在饲料中添加壳聚糖纳米颗粒可以提高尼罗罗非鱼的增重率和特定生长率以及体内淀粉酶和脂肪酶含量, 降低肠道细菌数量, 促进鱼生长。 田娟等用添加了壳寡糖质量分数为0、 0.10%、 0.30%、0.50%、 0.70%的饲料饲喂体质量为(3.02±0.16) g的吉富罗非鱼幼鱼。 结果发现在肠道主要菌群方面, 各壳寡糖添加组大肠杆菌数量均显著降低,乳酸杆菌数量显著增加; 在肠道组织结构方面,各壳寡糖添加组的幼鱼前肠组织均较对照组发育良好, 肠绒毛排列紧密, 高度和宽度均有增加。黄鑫玮等在饲料中添加壳寡糖投喂幼建鲤, 研究发现幼建鲤摄食含壳寡糖的饲料后的增重率和特定生长率均得到提高, 并且幼鱼肠道组织结构和菌落分布均发生了变化。 摄食了含壳寡糖饲料的幼鱼肠道的有益菌落双歧杆菌和乳酸杆菌的数量比对照组的有所增加, 且肠道内皱襞高度、 密度以及皱襞的肥大细胞和杯状细胞数量都有所增加。

甲壳素及其衍生物在虾类饲料中的添加, 可以提高虾的免疫能力, 促进虾的生长。
任季芳等做了两次实验, 一次是用0、0.25%、 0.50%、 0.75%壳聚糖溶液浸泡克氏原螯虾仔虾, 结果发现壳聚糖能提高仔虾体内ACP、AKP 活性, 并降低MDA 浓度; 另一次等在基础饲料中分别添加质量分数为0、 0.5%、 1.0%、1.5%、 2.0%、 3.0%的壳聚糖制成6 种试验饲料,投喂体质量为 (21.55±1.62) g 的克氏原螯虾60d。 试验测定了特定生长率、 死亡率、 退壳死亡率、 碱性磷酸酶、 谷丙转氨酶、 谷草转氨酶、肌肉营养成分、 肠道消化酶活性。 结果表明, 在成虾饲料中添加0.5%~1.5%的壳聚糖, 对克氏原螯虾具有一定的免疫保护和增强其消化机能的作用。 庄承纪等在做壳多糖对罗氏沼虾、 斑节对虾苗生长和抗菌防病作用研究, 结果发现在培养水体中加入壳多糖能净化水质,防止水质恶化; 壳多糖可抑制气单胞菌和弧菌的生长繁殖,增强抗病能力,提高虾苗的成活率。 胡琳琳等在基础饲料中分别添加0、 0.04‰、 0.15‰、 0.50‰的壳聚糖硫酸酯投喂凡纳滨对虾。 结果表明, 在饲料中添加壳聚糖硫酸酯可以明显提高对虾抵御白斑综合征病毒感染的能力。
陈伟军等在基础饲料中分别添加0、 0.2‰、0.4‰、 0.6‰、 0.8‰、 1.0‰、 2.0‰壳寡糖饲料,投喂体重均为(1.50±0.03) g 的凡纳滨对虾80d。结果表明, 饲料中添加壳寡糖能够增加凡纳滨对虾的肠道绒毛长度, 使上皮细胞排列得更加整齐、 紧密, 进而提高凡纳滨对虾的存活率和特定生长率, 起到促进凡纳滨对虾的生长。 Niu 等比较在饲料中添加甲壳素及其衍生物(壳聚糖、 壳寡糖及N-乙酰氨基葡萄糖) 对斑节对虾的影响,试验结果表明, 在饲料中添加壳聚糖的效果较佳, 可以提高斑节对虾的增重率、 存活率以及在氧胁迫下的耐受力。 马利等设置了含壳寡糖0、125mg/kg、 250mg/kg、 500mg/kg、 1000mg/kg 的5组饲料, 对初始体重约为1.23g 的南美白对虾进行8 周的饲养试验。 试验结果表明饲料中添加壳寡糖可以使南美白对血清中的高密度脂蛋白胆固醇显著增加, 促进脂类代谢。 Tseng 等从同形溞中提取甲壳素及其衍生物壳聚糖加入到饲料中投喂南美白对虾, 试验结果表明饲料中加入0.4%的壳聚糖可以改善虾的生长性能, 能提高其增重率和特定生长率。 Abdel-Warith 等用壳聚糖纳米颗粒和螺旋藻作为饲料添加剂投喂黑虎虾, 结果表明可以提高黑虎虾的特定生长率、 存活率。

李军涛在研究饲料中添加壳聚糖对方斑东风螺稚螺生长性能和免疫相关酶活性的影响中, 发现饲料中添加壳聚糖可以通过提高方斑东风螺稚螺肝脏中免疫相关酶的活性提高饲养成活率, 并能够在一定程度上加快生长速度。
张干等在基础饲料中分别添加0、 25、 50、100mg/kg 的低聚壳聚糖投喂均重为(16.20±1.30)g 的中华绒螯蟹56d, 结果表明, 饲料中添加50mg/kg 的低聚壳聚糖可以较好地提高中华绒螯蟹的生长性能、 非特异性免疫和抗氧化能力, 并降低机体脂肪沉积。 袁春营等在基础饲料中分别添加0.05‰、 0.1‰、 0.2‰、 0.4‰的低聚壳聚糖,对均重为(15.78±3.21) g 的中华绒螯蟹进行为期28d 的饲养试验。 通过测定中华绒螯蟹的血液免疫指标, 得出饲料中添加0.2‰的低聚壳聚糖可以显著提高中华绒螯蟹的免疫能力的结论。
魏文志等在基础饲料中分别添加0、 0.3%、0.6%、 1.2%的甲壳素, 对均重为(8.990±1.024g)的中华鳖进行为期31d 的饲养试验。 结果表明,添加0.6%的甲壳素能够显著促进中华鳖的生长和非特异性的免疫能力。
党慧凤等对仿刺参进行添加了壳寡糖的投喂试验。 试验结果表明饲料中添加了0.3%、 0.5%的壳寡糖可以作为免疫增强剂, 显著提高仿刺参各组织的ACP、 AKP、 SOD、 LZM 的活性。
4 小结
目前, 甲壳素及其衍生物在水产动物饲料上的应用越来越受到关注。 甲壳素在饲料中的添加可以提高水产动物生长性能和免疫能力。 而甲壳素的原料来源及加工工艺仍然有待进一步的研究和改善。 另外, 甲壳素在水产饲料中的适宜添加量也因水产动物的种类、 年龄、 体重等有所不同, 也是今后研究的重点。
猜你喜欢
中国果菜(2022年1期)2022-01-26 08:07:50
食品与机械(2020年7期)2020-01-05 02:54:03
中国化肥信息(2018年6期)2018-08-23 09:11:48
中学生数理化·高二版(2016年3期)2016-12-26 09:36:58
现代农业(2016年6期)2016-02-28 18:42:53
合成化学(2015年10期)2016-01-17 08:56:26
中国果菜(2015年2期)2015-03-11 20:01:05
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:58
无机化学学报(2014年10期)2014-02-28 17:33:13
河南科技(2014年7期)2014-02-27 14:11:10
