
生物质炭对铝胁迫下观光木幼苗生长生理特性的影响
2021-10-21 11:35潘陆荣亢亚超王凌晖
西南林业大学学报 2021年5期
潘陆荣 亢亚超 潘 虹 王凌晖
(广西大学林学院,广西 南宁 530004)
随着铝在人类生产生活中大量生产和使用,包括铝盐絮凝剂在水处理、湖泊治理中的广泛应用以及环境酸化,越来越多的铝进入生态系统,给许多生物带来毒害效应。大量研究表明铝毒是酸性土壤中抑制植物根伸长和植株生长,导致人工林产量下降和森林退化的主要原因之一[1-2],防治铝毒害的方法主要有物理、化学和生物方法等[3]。其中,施加改良剂是修复铝毒害土壤的一种较经济的,且在实际中获得应用的原位化学处理方法[4]。生物质炭高度稳定,离子交换能力强,能吸附环境中的重金属和有机污染物,因此成为了治理重金属污染和有机污染物污染的一种重要的改良剂[5]。目前,通过施加生物质炭来缓解重金属对植物毒害的研究也取得一定进展,如黄韡等[6]针对生物质炭对铝胁迫下常绿杨(Populus simoniivar.przewalskii)生理特征进行研究,结果表明生物质炭的用量为3%时,对铝胁迫状态下常绿杨的生长状况的改善作用最为明显;陆文龙等[7]针对生物质炭对镉胁迫下小白菜(Brassica rapavar.glabra)生长特性进行研究,结果表明生物质炭对小白菜株高和叶宽的指标都有不同程度的促进作用。在施加生物质炭并且受镉污染的小白菜的根系耐性指数大于1.0。
观光木(Tsoongiodendron odorum)属木兰科(Magnoliaceae)观光木属(Tsoongiodendron),别名香花木和宿轴木兰,为单属种[8],是我国珍稀Ⅱ级保护植物,观光木树形高大挺拔、材质优良、便于加工,既可用于园林绿化,也是优良的用材树种[9-10]。目前,有关观光木的研究主要包括观光木群落结构[11]、遗传育种及造林[12]、木材材质[13]以及土壤理化性状[14]、生理生化特征[15-16],其中涉及到观光木胁迫研究的包括干旱胁迫[17]、低温胁迫[18]、镉胁迫[19]、盐分胁迫[20]等。而施加生物质炭对铝胁迫下观光木幼苗生长生理特性的研究未见,中国南方分布着大面积富铝化酸性土壤,更容易产生活性铝抑制南方地区林木生长,观光木推广栽培过程将面临着铝毒害的现实。因此,研究观光木耐铝机制,并通过施加外源物质缓解铝对观光木的毒害作用,是充分利用酸性土地资源的有效途径之一。本研究通过设置不同生物质炭浓度和铝浓度,比较不同处理对观光木幼苗生长生理特性的影响,研究生物质炭对铝胁迫下观光木幼苗生长生理方面的缓解作用,以期为指导观光木种植,提高酸性土壤利用率提供参考依据。
1 材料与方法
1.1 试验材料
试验材料为1年生观光木幼苗,2019年5月底将观光木幼苗移植到广西大学林学院苗圃教学基地(108°17′E,22°51′N)中,采用规格为17 cm(径)×14 cm(高)的有孔塑料盆,培养基质由体积比为3∶1的土壤和河沙混合,试验前测定土样养分含量为全氮1.096 g/kg,全磷0.610 g/kg,全钾5.090 g/kg,有机质11.730 g/kg,pH值4.9。每盆1株观光木幼苗,每日浇水,移植1周后向各盆中浇1 L改良霍格兰营养液,缓苗1个月后开始处理。铝的供体为分析纯AlCl3-6H2O4,生物质炭由观光木的枯枝落叶在高温缺氧条件下热解制成,具体制备参考Lu等[21]的方法。
1.2 试验设计
试验采用完全随机设计方法,参考杨梅等[22]和杨林通[23]的研究,设置3个生物质炭浓度和2个铝浓度,共6个处理(表1),每个处理9株盆栽。每2周浇1次霍格兰营养液,每周浇1次100 mL处理液并施加生物质炭,处理液的pH使用HCl调至4.1~4.2,确保为酸性环境。共处理12周。

表1 生物质炭铝互作试验设置Table 1 The experimental design of the interaction between biochar and aluminum
1.3 测定指标及方法
1.3.1 生长指标的测定
试验开始即使用皮尺(精确到0.1 cm)测定幼苗株高,用数显游标卡尺(精确到0.01 mm)测定地径,试验期间每月测定1次,每组每株重复测定3次。2019年10月试验结束后,每个处理组挖取3株观光木幼苗,采用烘干称质量法[24]测定生物量。使用EXPRESSION 1 680根系扫描仪(爱普生,日本)测定各项根系指标,每个处理选取3株观光木幼苗破坏取样,测定其根系的总根长、根尖数、根总体积、根总表面积和根平均直径。
1.3.2 生理指标的测定
2019年10月试验结束时,每个处理采集多片完整叶片,用冰盒保存带回实验室进行生理指标的测定,生理指标测定参考《植物生理学实验指导》[25],其中每个处理做3次重复。采用硫代巴比妥酸(TBA)法测定丙二醛(MDA)含量;蒽酮比色法测定叶片可溶性糖含量;考马斯亮蓝法测定叶片可溶性蛋白含量;酸性茚三酮法测定游离脯氨酸含量;丙酮-乙醇混合提取法测定叶绿素含量;根茎叶铝含量的测定则采用李合生[26]的干灰化法进行测定。
1.4 数据分析
试验数据采用Excel2010进行统计分析,并绘制图表,采用DPS软件进行方差分析,多重比较采用LSD方法进行比较,应用模糊综合评价法[27-28]根据隶属度理论把定性评价转化为定量评价,对所有测定的生长和生理指标进行综合评价。隶属函数的计算公式如下。
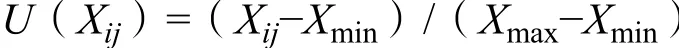
式中:U(Xi)表示i处理j指标的耐铝隶属函数值;Xij表示i处理j指标的测定值,Xmax、Xmin分别代表各处理中指标的最大值和最小值。
2 结果与分析
2.1 生物质炭铝互作对观光木幼苗生长指标的影响
2.1.1 生物质炭铝互作对观光木幼苗株高及地茎的影响
由图1可知,当生物质炭施加量一定时,铝胁迫组株高增量、地径增量均低于未施加铝组;C1Al2的株高增量、地径增量较C1Al1分别减少了18.37%、27.66%;C2Al2的株高增量、地径增量较C2Al1分别减少了15.24%、18.39%;C3Al2的株高增量、地径增量极显著低于C3Al1(P<0.01),较C3Al1分别减少了17.53%、33.93%;说明施加铝在一定程度上降低了观光木幼苗的株高增量、地径增量。无铝胁迫施加的情况下,C2Al1、C3Al1的株高增量、地径增量均高于C1Al1,且C3Al1的生物质炭株高增量、地径增量均极显著高于C1Al1和C2Al1(P<0.01),说明施加生物质炭有利于观光木幼苗株高增量、地径增量的增加,尤其是C3Al1的生物质炭增加量最大。
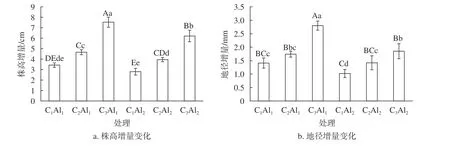
图1 生物质炭铝互作对观光木幼苗株高及地茎的影响Fig. 1 Effect of biochar-aluminum interaction on plant height and ground diameter of T. odorum seedlings
铝胁迫环境下,C2Al2、C3Al2的株高增量、地径增量均高于C1Al2,C2Al2的株高增量、地径增量较C1Al2分别提高了29.11%、28.17%;C3Al2的株高增量、地径增量较C1Al2分别提高了54.91%、44.86%;说明施加生物质炭能够有效缓解铝对观光木幼苗胁迫作用,同样是C3Al2的生物质炭增加量最大。
2.1.2对观光木幼苗生物量的影响
由图2可知,当生物质炭施加量一定时,铝胁迫组根生物量、茎生物量、叶生物量均低于未施加铝组;C1Al2的根、茎、叶生物量较C1Al1分别减少了31.51%、32.77%、28.53%;C2Al2的根、茎、叶生物量较C2Al1分别减少了36.27%、22.48%、26.93%;C3Al2的根、茎、叶生物量较C3Al1分别减少了23.93%、24.03%、24.89%;说明施加铝抑制观光木幼苗生长,从而降低其根茎叶生物量。无铝胁迫施加的情况下,C2Al1、C3Al1的根生物量、茎生物量、叶生物量均极显著高于C1Al1(P<0.01),C2Al1的根、茎、叶生物量较C1Al1分别提高了40.87%、23.45%、45.37%;C3Al1组根、茎、叶生物量较C1Al1分别提高了64.84%、40.95%、66.29%;说明施加生物质炭有利于观光木幼苗根、茎、叶生物量的积累,且C3Al1的生物质炭积累效果最佳。

图2 生物质炭铝互作对观光木幼苗生物量的影响Fig. 2 Effect of biochar-aluminum interaction on biomass of T. odorum seedlings
铝胁迫环境下,C2Al2、C3Al2的根生物量、茎生物量、叶生物量均高于C1Al2,C2Al2根茎叶生物量较C1Al2分别提高了36.45%、33.61%、46.56%;C3Al2的根茎叶生物量较C1Al2分别提高了68.35%、47.80%、67.93%;说明施加生物质炭能显著改善铝对观光木幼苗生长的抑制作用。
2.1.3 对观光木幼苗根系的影响
由表2可知,当生物质炭施加量一定时,铝胁迫组的总根表面积、根平均直径、总根长、根尖数、总根体积均低于未施加铝组;C1Al2的各项根系指标较C1Al1分别减少了18.44%、54.46%、67.89%、14.23%、31.73%;C2Al2的各项根系指标较C2Al1分别减少了13.33%、40.98%、48.63%、9.11%、25.53%;C3Al2的各项根系指标较C3Al1分别减少了10.57%、28.97%、32.97%、14.59%、23.67%;说明施加铝对观光木幼苗生长产生胁迫,抑制了观光木幼苗的根系的生长。无铝胁迫施加的情况下,C2Al1、C3Al1的总根表面积、根平均直径、总根体积、总根长、根尖数均高于C1Al1,C2Al1总根表面积、根平均直径、总根体积、总根长、根尖数较C1Al1分别提高了14.97%、18.05%、24.90%、6.67%、8.80%;C3Al1的各项根系指标较C1Al1分别提高了32.92%、32.09%、48.24%、20.41%、24.36%;说明施加生物质炭有利于促进观光木幼苗根系的生长,尤其是C3Al1的促进效果最佳。

表2 生物质炭铝互作对观光木幼苗根系的影响Table 2 Effect of biochar-aluminum interaction on root of T. odorum seedlings
铝胁迫情况下,C2Al2、C3Al2的总根表面积、根平均直径、总根体积、总根长、根尖数均高于C1Al2,C2Al2较C1Al2分别提高了19.99%、31.21%、53.05%、11.93%、16.40%;C3Al2较C1Al2分别提高了38.83%、52.63%、75.20%、20.07%、32.35%;说明施加生物质炭能够有效缓解铝对观光木幼苗胁迫作用,促进观光木幼苗根系的生长。
2.2 生物质炭铝互作对观光木幼苗生理特性的影响
2.2.1 对观光木幼苗叶片MDA含量的影响
由表3可知,施加相同生物质炭量的情况下,铝胁迫组叶片MDA含量均高于其对应的未施加铝组,增加量分别为未施加铝组的21.19%、22.96%和31.76%。可见,铝胁迫下观光木幼苗受到了严重伤害,叶片过氧化水平显著提高(P<0.05);同时也提高了叶片MDA含量。未施加铝处理时,C2Al1、C3Al1的叶片MDA含量均低于C1Al1,其减少量分别为C1Al1的5.85%和14.50%,说明施加生物质炭有利于降低叶片MDA含量;铝胁迫条件下,MDA含量随生物质炭的增加而表现出连续下降的趋势,C2Al2、C3Al2的减少量分别为C1Al2的4.48%和7.04%,两者之间差异不显著;说明铝胁迫环境下,施加生物质炭能够缓解铝毒害,有效降低叶片MDA含量,增加植株耐铝性,且C2Al2、C3Al2之间缓解效果差异不显著。

表3 生物质炭铝互作对观光木幼苗抗性生理指标的影响Table 3 Effect of biochar-aluminum interaction on physiological index of leaf resistance of T. odorum seedlings
2.2.2 对观光木幼苗叶片可溶性糖含量的影响
由表3可知,施加相同生物质炭量的情况下,铝胁迫组叶片可溶性糖含量均低于其对应的未施加铝组,减少量分别为未施加铝组的16.27%、15.09%和12.12%。可见,铝胁迫能抑制观光木叶片可溶性糖的含量,而施加生物质炭能够有效缓解铝毒害,增加植株耐铝性,且C3Al2的缓解效果最好。未施加铝处理时,C2Al1、C3Al1的叶片可溶性糖的含量均高于C1Al1,与C1Al1相比,C2Al1、C3Al1叶片可溶性糖含量分别增加了7.84%和29.41%,施加生物质炭有利于提高叶片可溶性糖含量;铝胁迫条件下,C2Al2、C3Al2分别为C1Al2的1.09倍、1.36倍;说明在铝胁迫环境下,施加生物质炭有效增加观光木叶片可溶性糖含量,且与生物质炭施加量呈正相关。
2.2.3对观光木幼苗叶片可溶性蛋白的影响
由表3可知,施加相同生物质炭量的情况下,铝胁迫组叶片可溶性蛋白含量均低于其对应的未施加铝组,减少量分别为其未施加铝组的10.03%、9.40%和7.48%。可见,铝胁迫能抑制观光木叶片可溶性蛋白的含量;未施加铝处理时,与C1Al1相比,C2Al1、C3Al1的叶片可溶性蛋白含量均有所增加,但无显著性差异,其增加量分别为C1Al1的6.18%和8.56%;铝胁迫条件下,叶片可溶性蛋白的含量随生物质炭的增加而持续增加,C2Al2、C3Al2的增加量分别为C1Al2的7.34%和12.13%;说明施加生物质炭能够在一定程度上增加叶片可溶性蛋白含量,增强植物耐铝性。
2.2.4 对观光木幼苗叶片游离脯氨酸含量的影响
由表3可知,施加相同生物质炭量的情况下,其对应铝胁迫组叶片脯氨酸含量均有增加,增加量分别为对应未施加铝组的27.43%、22.14%和70.08%。可见,铝胁迫对叶片渗透势的影响是极显著(P<0.01)的;未施加铝处理时,C2Al1、C3Al1的叶片脯氨酸含量均低于C1Al1,其减少量分别为C1Al1的9.26%和21.98%,且C3Al1的叶片脯氨酸含量极显著(P<0.01)低于C1Al1;铝胁迫环境下,C2Al2、C3Al2的叶片脯氨酸含量均小于C1Al2,其减少量分别为C1Al2的13.03%和17.78%;说明添加生物质炭能减少铝胁迫对叶片渗透势的伤害,减少脯氨酸的产生,且C3Al2的缓解效果较好。
2.2.5 对观光木幼苗叶片光合色素的影响
由图3可知,当生物质炭施加量一定时,铝胁迫组叶片叶绿素a、叶绿素b含量均有所减少。铝胁迫环境下,叶片叶绿素a、叶绿素b含量随生物质炭施加量的增加而增加,但C2Al2与C1Al2无显著差异,C3Al2的叶绿素a、叶绿素b含量极显著增加(P<0.01);说明铝胁迫能抑制光合色素含量,未施加铝处理时,叶片叶绿素a含量随着生物质炭施加量的增加而极显著增加(P<0.01),分别增加至0.457、0.615 mg/g,C3Al1的叶绿素b含量与C1Al1存在极显著性差异(P<0.01),其含量为0.341 mg/g,C3Al1的叶片光合色素含量极显著增加(P<0.01);铝胁迫条件下,C3Al2能有效缓解胁迫对叶绿素a和叶绿素b含量的抑制作用,叶片光合色素含量极显著增加(P<0.01)。

图3 生物质炭铝互作对观光木幼苗叶片光合色素的影响Fig. 3 Effect of biochar-aluminum interaction on photosynthetic pigment of T. odorum seedlings
2.3 生物质炭铝互作下观光木幼苗不同部位中铝含量分布
由图4可知,未施加生物质炭的情况下,铝胁迫组根茎叶铝含量均有不同程度增加,其增加量分别为32.86%、10.61%、1.71%。可见,铝胁迫对根部和叶片铝含量的影响是极显著的(P<0.01);未施加铝处理时,C2Al1、C3Al1根茎叶铝含量均低于C1Al1,且C3Al1的根茎叶铝含量与C1Al1存在极显著性差异(P<0.01);铝胁迫环境下,根部、茎部和叶片铝含量随生物质炭施加量的增加而持续减少,C2Al2的茎部和叶片铝含量极显著低于C1Al2(P<0.01),而根部铝含量与C1Al2无显著差异;C3Al2的根部、茎部和叶片铝含量均极显著低于C1Al2(P<0.01),减少量分别为12.13%、21.32%、16.23%;说明施加生物质炭对缓解观光木根、茎、叶铝胁迫有极显著作用(P<0.01)。

图4 生物质炭铝互作下观光木幼苗不同部位中铝含量Fig. 4 Aluminum content in different parts of T. odorum seedlings under the interaction of biochar-aluminum
2.4 生物质炭铝互作下观光木各指标的隶属函数分析
由表4可知,各处理的隶属函数均值从大到小 依 次 为C3Al1> C3Al2> C2Al1> C1Al1> C2Al2>C1Al2。未施加生物质炭时,C1Al2隶属函数值排最后。可见,施加铝明显抑制观光木幼苗生长,而铝胁迫环境下,C3Al2的隶属函数值大于C2Al2和C1Al2。说明C3Al2的缓解铝胁迫效果最佳。
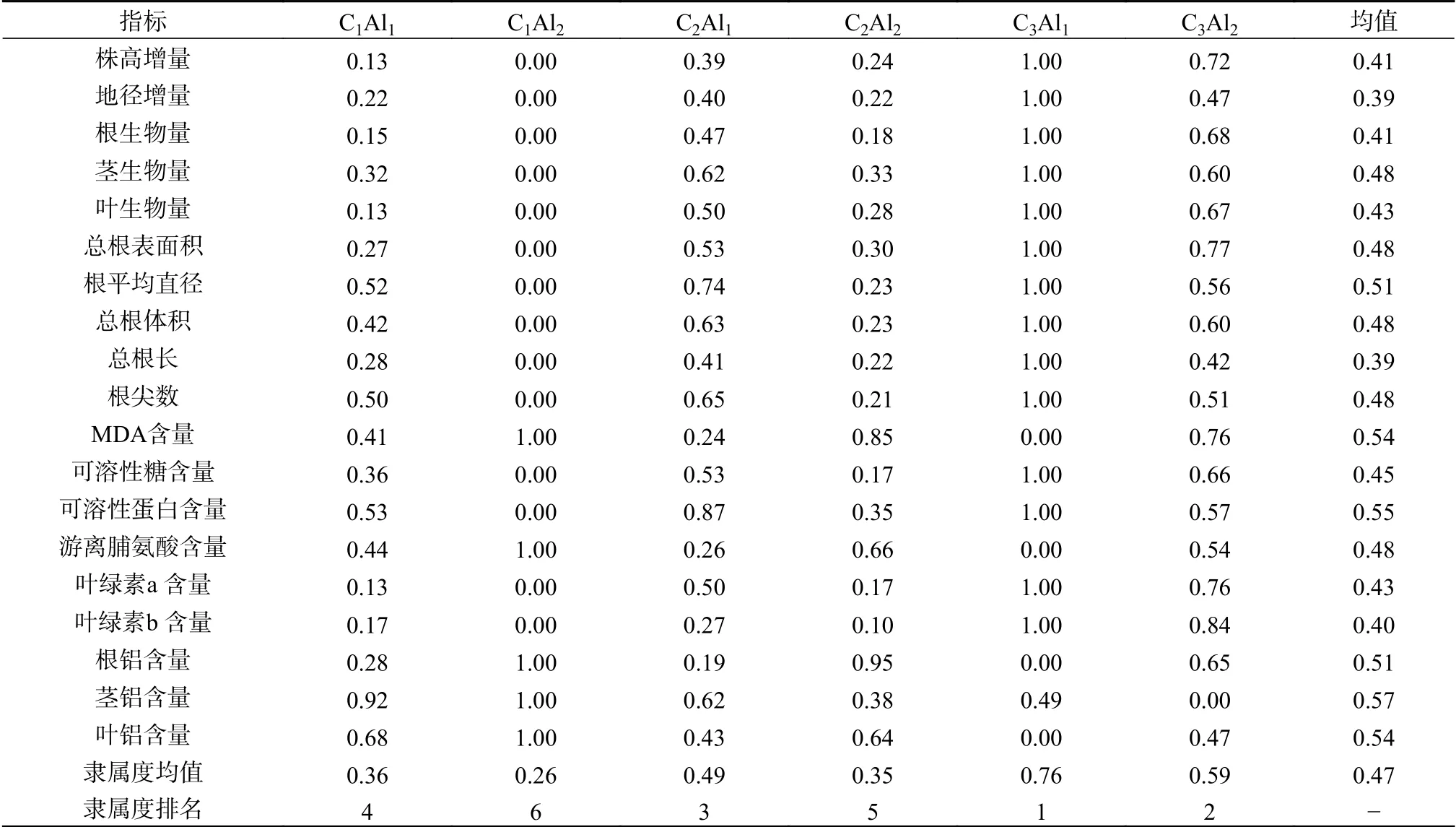
表4 生物质炭铝互作下观光木各指标的隶属函数分析Table 4 Membership function analysis of T. odorum seedlings under the interaction of biochar-aluminum
3 结论与讨论
当植物处于胁迫环境时,其外在形态和生理生化过程会受到不同程度的影响,植物根伸长和植株生长受到抑制,细胞膜透性增大、抗氧化酶提高以及光合作用受损等,造成生长发育不良、植株矮化的现象[29]。在本研究中,当生物质炭施加量一定时,铝胁迫组株高增量、地径增量、根生物量、茎生物量和叶生物量均低于未施加铝组,观光木幼苗生长受到抑制,这与前人关于铝胁迫能显著抑制植物生长的结论相一致[30]。在本研究中,无铝胁迫施加的情况下,施加生物质炭有利于观光木幼苗株高增量、地径增量以及生物量的增加,尤其是施加8 g生物质炭时增加量最大,有研究表明,施用生物质炭可降低土壤的酸度和重金属的生物有效性,促进植株生长、增加作物产量[31]。在本研究中,铝胁迫环境下,施加生物质炭组的株高增量、地径增量以及生物量均高于未施加生物质炭组,说明施加生物质炭能够有效缓解铝对观光木幼苗胁迫作用,这与牛玉德等[32]针对不同生物质炭施用量对汉中烤烟生长发育的研究结果相似。铝胁迫处理不但显著抑制观光木株高、地径、生物量等指标的增长,对其根系也具有明显的影响,普遍认为铝对植物的毒害首先表现在植物根部,植物根系的伸长以及侧根数量的增加都会受到抑制,导致根尖短粗、根冠脱落[33]。在本研究中,当生物质炭施加量一定时,铝胁迫组的总根表面积、根平均直径、总根长、根尖数、总根体积均低于未施加铝组,观光木幼苗根系生长受到显著抑制;无铝胁迫施加的情况下,施加生物质炭有利于促进观光木幼苗根系的生长,尤其是施加8 g生物质炭时促进效果最佳;在铝胁迫情况下,施加生物质炭组的各项根系指标均高于未施加生物质炭组,说明施加生物质炭能够有效缓解铝对观光木幼苗胁迫作用,促进观光木幼苗根系的生长,这与王欢欢等[34]研究结果相似,值得注意的是,许唯等[35]研究表明,不同生物质炭均抑制了玉米(Zea mays)幼苗主根的伸长,对玉米根系的生长发育存在一定毒害作用。因此,还需针对特种植物生长及土壤相关生物学性质进一步研究分析。
逆境胁迫下植物易产生超氧化物自由基,由于不饱和脂肪酸发生了氧化反应,膜脂过氧反应的产物MDA由此得到积累,质膜受到损伤,其选择透过性变差导致细胞内电解质外渗,细胞内环境的相对平衡被打破。而脯氨酸是植物体内最重要和有效的渗透调节物质,在逆境胁迫下会迅速积累,通过提高细胞液浓度降低其渗透势,以缓解逆境胁迫[36]。可溶性蛋白能提高植物细胞的保水能力,对细胞的生命物质及生物膜起保护作用,而可溶性糖是植物生长发育和基因表达的重要调节因子,也是一类渗透调节物质[37]。在本研究中,当生物质炭施加量一定时,铝胁迫组观光木叶片膜脂过氧化的产物MDA和游离脯氨酸含量均增大,细胞膜透性增大,细胞可溶性糖、可溶性蛋白的含量降低,与柑橘(Citrus reticulata)遭受胁迫时生理上表现的结果相似[38];此外,观光木根、茎、叶铝含量也增加,而叶片中的叶绿素a、叶绿素b含量积累随之减少,可能是因为铝与叶绿素合成的叶绿素酸酯还原酶等部分结合,降低叶绿素酸酯还原酶酶活性[39]。叶绿素含量的下降不利于植物叶片捕捉光合作用需要的光能,严重影响叶片光合作用的正常进行。未施加铝胁迫或施加铝胁迫情况下,随着生物质炭的施加,观光木幼苗叶片中可溶性糖和可溶性蛋白、叶绿素a、叶绿素b、根茎叶铝含量等均有不同程度增加,而MDA和游离脯氨酸含量随之降低,施加生物质炭对促进观光木幼苗生长、缓解铝胁迫均有一定效果,也是施加8 g生物质炭时效果最佳。
由隶属函数分析可知,各处理的隶属函数均值从大到小依次为C3Al1> C3Al2> C2Al1> C1Al1>C2Al2> C1Al2,施加铝明显抑制观光木幼苗生长,而铝胁迫环境下,C3Al2的隶属函数值大于C2Al2和C1Al2。综上所述,表明在本试验条件下,施加生物质炭能够促进观光木幼苗生长,有效缓解铝胁迫对观光木生长、光合色素以及抗性生理造成的伤害,提高观光木的耐铝性,且施加8 g生物质炭时,缓解铝胁迫效果最佳,为指导观光木种植,提高酸性土壤利用率提供参考依据。
猜你喜欢
今日农业(2022年1期)2022-11-16
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
北京航空航天大学学报(2022年5期)2022-06-06
财会月刊·下半月(2022年4期)2022-04-25
当代陕西(2022年6期)2022-04-19
南京信息工程大学学报(2022年1期)2022-04-09
今日农业(2021年21期)2022-01-12
小学生(看图说画)(2020年9期)2020-09-17
