
沙质草地植物功能性状对放牧、增水、氮添加及其耦合效应的响应机制
2021-10-11 07:23:44左小安
生态学报 2021年18期
张 晶,左小安
中国科学院西北生态环境资源研究院,兰州 730030
放牧引起的草地退化问题早已备受关注,尤其是在我国,过度放牧导致的草地生产力持续衰减、生物多样性丧失和生态系统服务功能下降等问题已经成为当前草地生产及理论方面的棘手难题[1]。季节性放牧是传统的草原利用方式。研究通过比较春季、夏季和秋季放牧对草原植物生长和动物增重的影响,发现非生长季放牧能够在不损害可再生自然资源(植物生产)的情况下最大化动物生产[2],是实现草地科学管理和可持续发展的有效方式。全球气候模型预测,降水格局会随着全球变暖而发生改变[3-4],主要表现为:降水总量、降水分布发生了改变,以降水间隔时间增长、单次降水量增大为特征的极端降水事件的频率将会持续增加[2-3,5]。大量研究表明,比起年降水量,生长季降水在调控草地植被生长过程方面发挥着重要的作用[6],植物功能性状指植物对外界环境长期适应与进化后所表现出的可量度、与生产力优化或环境适应等密切相关的属性[7]。从性状角度出发能够将植物的适应策略、群落构建机制和生态过程等有机结合,揭示环境变化对群落生产力的影响机制[8-9],因此通过监测生长季降水格局对草地植物功能性状的影响可推演出草地生态系统功能对极端降水事件的响应。伴随着工农业活动的加剧,通过大气干湿沉降输入到陆地生态系统中的含氮化合物日益增多[10]。氮沉降是导致生物多样性和生态系统稳定性丧失的主要环境影响因子,同时氮肥也被用作提高生产力的主要营养元素来促进退化草地的恢复[11]。因此,氮素对不同区域和生态系统的作用是不一致、甚至是截然相反的。
植物功能性状是指影响植物的生存和适应以及影响植物获取、利用和贮存资源的属性,可以全面地反映有关植物生长和对环境适应的重要信息[12-14]。总体而言,功能性状包括了有关于植物形态、生理、物候特性等各个方面[15-16],通过研究植物功能性状变化不仅能解释植物对环境变化的适应机理[17],还能预测植物群落的分布格局和组成[13],准确评估生态系统功能[18]。在我国半干旱草原区,放牧是草地利用和管理的重要方式,水分和氮素是限制植物生长最重要的两种资源,因此,在人类活动和气候变化背景下深入揭示放牧、降水变化和氮添加及其交互作用下本区域植物功能性状的变化规律,不仅有助于预测未来草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学意义。
科尔沁沙地是我国北方半干旱地区农牧交错带的典型区域,近半个世纪以来由于过度放牧、降水格局变化、农业活动引起的氮沉降的影响,导致该区域生物资源锐减和生产力下降,成为我国沙漠化最严重的地区之一[19]。在该区域,生长季降水在调控草地植被生长过程及生物量积累方面发挥着主要的作用[20],因此,深入研究全球变化和人类活动对该区域植物的功能性状的影响及调控机制,能够对认识半干旱沙质草地植被生长过程中的适应对策,预测未来沙质草地生态系统功能变化提供充足的数据支撑和科学的管理依据。
1 数据与方法
1.1 研究区概况
本研究在中国科学院奈曼沙漠化研究站附近的沙质草地上进行。草地中以一年生(狗尾草Setariaviridis(L.)Beauv、蒺藜TribulusterretrisL、尖头叶藜ChenopodiumacuminatumWilld)和多年生(糙隐子Cleistogenessquarrosa(Trin.)Keng、胡枝子LespedezabicolorTurcz、白草PennisetumcentrasiaticumTzvel)草本植物为主(表1)。该区气候属典型的半干旱大陆性季风气候,年平均降水量为360 mm,其中近75% 集中在生长季节(5月至8月)。土壤为砂质土壤,参照中国土壤分类系统(http://www.resdc.cn)。

表1 沙质草地主要优势植物生活型和功能群特征Table 1 The characteristics of life form and functional group of dominant plants in sandy grassland
1.2 试验设计
2014到2016年间,选取异质性较小的同一块退化沙质草地开展封育/放牧、氮添加和增水的三因素控制实验,样地面积约为0.67 hm2,植被平均盖度为60%—90%(2015年8月调查数据)。放牧方式为非生长季(9月到第二年3月)放牧,生长季(4月到8月)禁牧。放牧强度为1头牛/hm2,属于重度放牧[21];氮添加方式为5月和7月各添加10 g N/m2,总计20 g N/m2,氮肥以尿素的形式进行添加;每年按月降水量的30%进行增水。每种措施6个重复,总计48个样地,每个样地为8 m×8 m,样地间距为1 m。
2016年8月对每个样地内优势物种(每个样地4—6种植物)叶片的功能性状进行取样和测定。植物功能性状的指标主要包括:植物高度(height)、比叶面积(SLA)、叶片干物质含量(LDMC)、叶片氮含量(LNC)和叶片水分利用效率(δ13C)。在每个样地内针对每种优势植物各取植株5—10株,选择10片无病虫害的叶片展开,夹在湿润的滤纸间,迅速放入密封袋贮藏于铺满冰袋的便携式冰箱中(<5℃)。回到实验室将叶片浸入纯净水中,放置于黑暗的环境中(5℃)冷藏12 h。叶片取出后迅速用滤纸吸干表面的水分,在百万分之一的电子天平上称取叶鲜重。随后将叶片平铺在扫描仪上扫描,再用ImageJ 1.48软件进行处理,得到叶面积。最后,将扫描后的叶片放入60℃烘箱内烘干48 h,取出称得叶干重。LDMC=叶干重/叶鲜重,SLA=叶面积/叶干重。为了保证每个植物中都有足够的叶片用来测量样品的氮含量和水分利用效率,在每个采样植株附近获取足够的植物叶片样品,与原来采集烘干的植物样品相混合粉碎,过0.25 mm筛,存储备用。分析前在65℃的干燥箱中烘干1 h,放入干燥器中保存待测。采用元素分析仪(Costech ECS 4010,意大利)测定样地中每种植物的叶片LNC。叶片的碳同位素采用元素分析仪和碳同位素分析仪(Picarro 2101,美国)测定,δ13C值采用PDB标准,分析误差≤0.15‰。水分利用效率根据碳同位素比值及经验公式计算,降水利用效率为生物量与降水量的比值。
1.3 数据分析
植物功能多样性分析方法[22]:
植物群落功能性状(CWM),即就某一性状而言,群落内所有该性状的值与其相对生物量积的总和,表示群落内某一功能性状的均值,常用来评估群落动态和生态系统特征,CWM由下列公式计算:
(1)
式中,Pi是物种i在群落内的相对生物量,n是群落内物种总数,traiti是物种i的性状值。
1.4 统计分析
采用SPSS 21.0进行数据分析,所有数值均采用平均值±标准误表示。三因素方差分析比较放牧、增水和氮添加对沙质草地植物群落(CWM)、一年生和多年生植物功能性状的影响;多重比较采用最小显著性差异法(LSD),主成分分析法(PCA)检验植物功能性状之间的相关关系。
2 结果与分析
2.1 植物群落功能性状对放牧、增水、氮添加及其耦合效应的响应
本研究中,氮添加对CWMheight、CWMLDMC和CWMLNC具有极显著影响(P<0.01),放牧和增水的耦合效应对CWMSLA具有显著影响(P<0.05),而单因素的放牧和增水对植物群落功能性状无显著影响(表2)。

表2 植物群落功能性状对试验处理响应的方差分析Table 2 Analysis of variance (ANOVA) of plant community functional traits (CWM) in responses to experimental treatments
CWMheight和CWMLNC在氮添加处理下分别显著提高了36.8%和73.6%(P<0.05),而CWMLDMC在氮添加处理下显著降低了47.7%(P<0.05)。在封育情况下,CWMSLA在增水处理下显著降低了14.3%(P<0.05),而在放牧情况下,增水对CWMSLA无显著的影响;在正常降水情况下,CWMSLA在放牧处理下显著降低了12.1%(P<0.05),而在增水情况下,放牧对CWMSLA无显著影响(图1)。
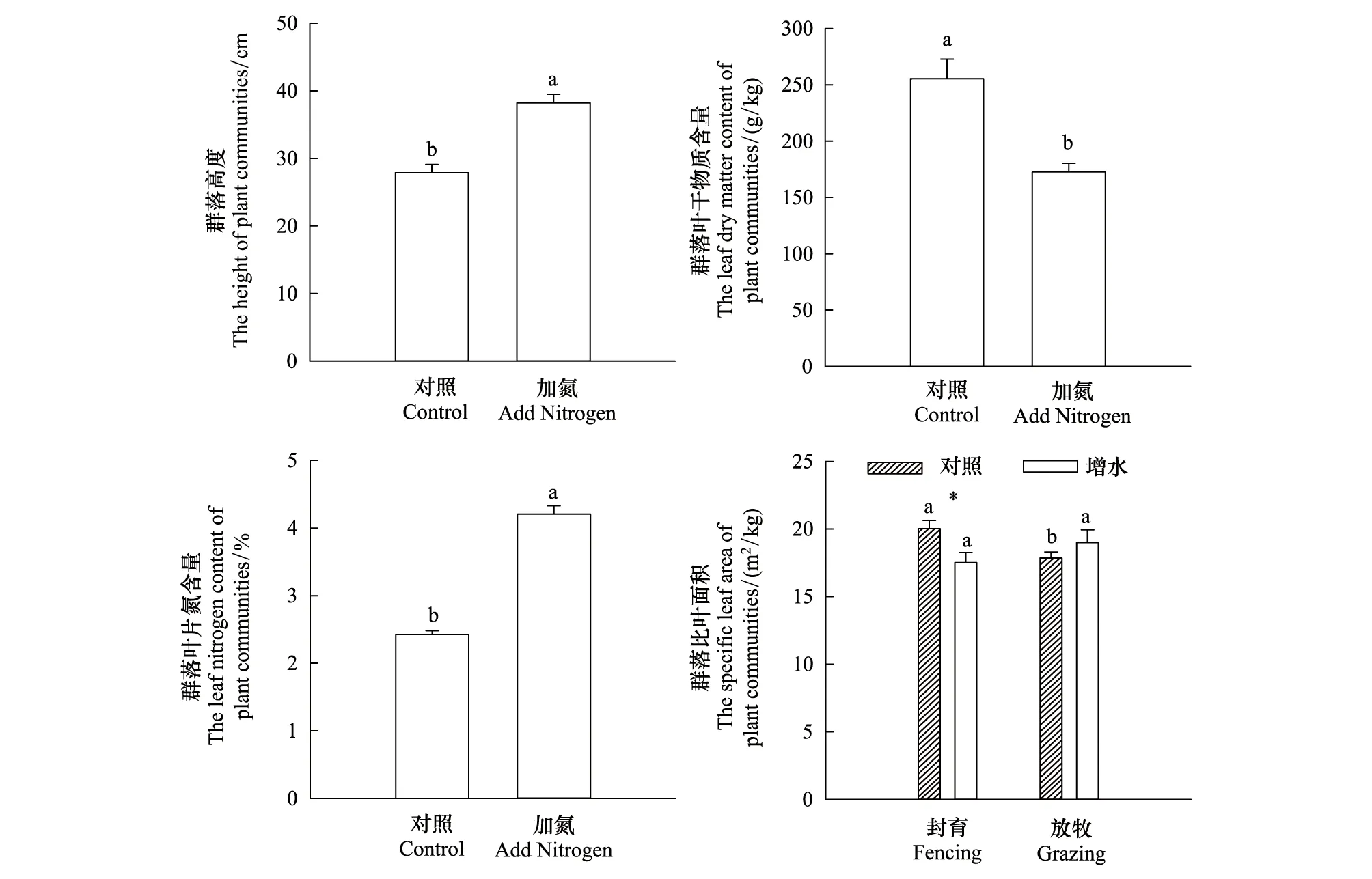
图1 氮添加、放牧和增水耦合效应对植物群落功能性状(CWM)的影响Fig.1 Effects of nitrogen addition, coupling effects of grazing and increase water on plant community functional traits (CWM)不同小写字母表示不同处理下植物群落功能性状在P<0.05水平上的差异性;* 表示封育管理条件下增水处理对植物群落功能性状在P<0.05水平上的差异性
2.2 一年生植物平均功能性状对放牧、增水、氮添加及其耦合效应的响应
本研究中,放牧对一年生植物的高度具有显著影响(P<0.05),氮添加对一年生植物的高度、LNC和δ13C具有极显著影响(P<0.01),单因素的增水对一年生植物功能性状无显著影响(表3)。
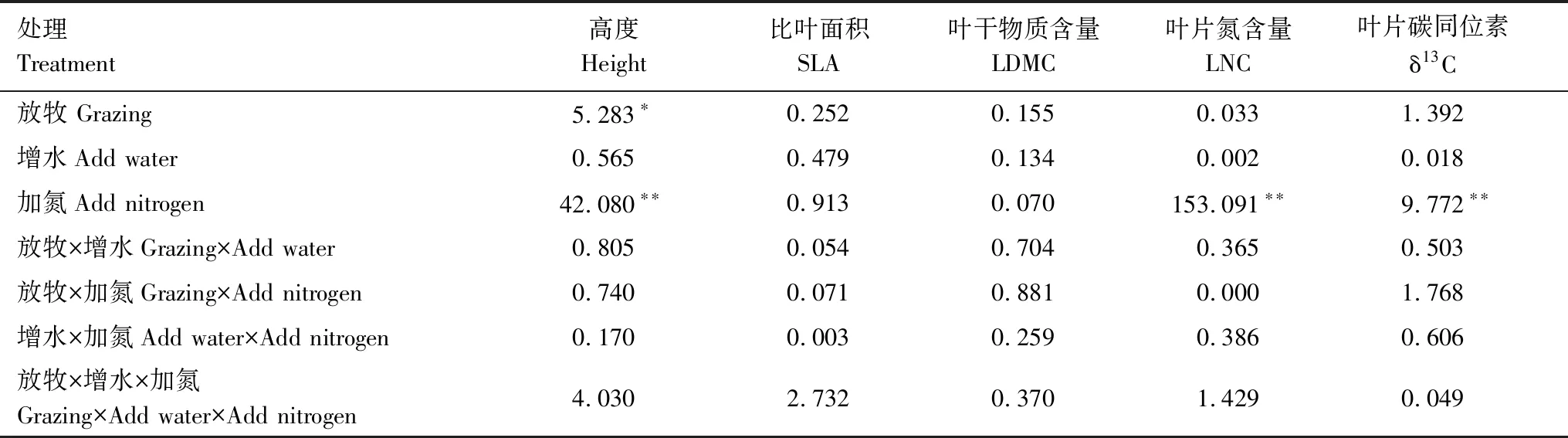
表3 一年生植物平均功能性状对试验处理响应的方差分析Table 3 Analysis of variance (ANOVA)of annuals in responses to experimental treatments
一年生植物高度在放牧处理下显著降低了12.5%(P<0.05),一年生植物高度、LNC和δ13C在氮添加处理下分别显著提高了39.7%、68.9%和12.2%(P<0.05)(图2)。
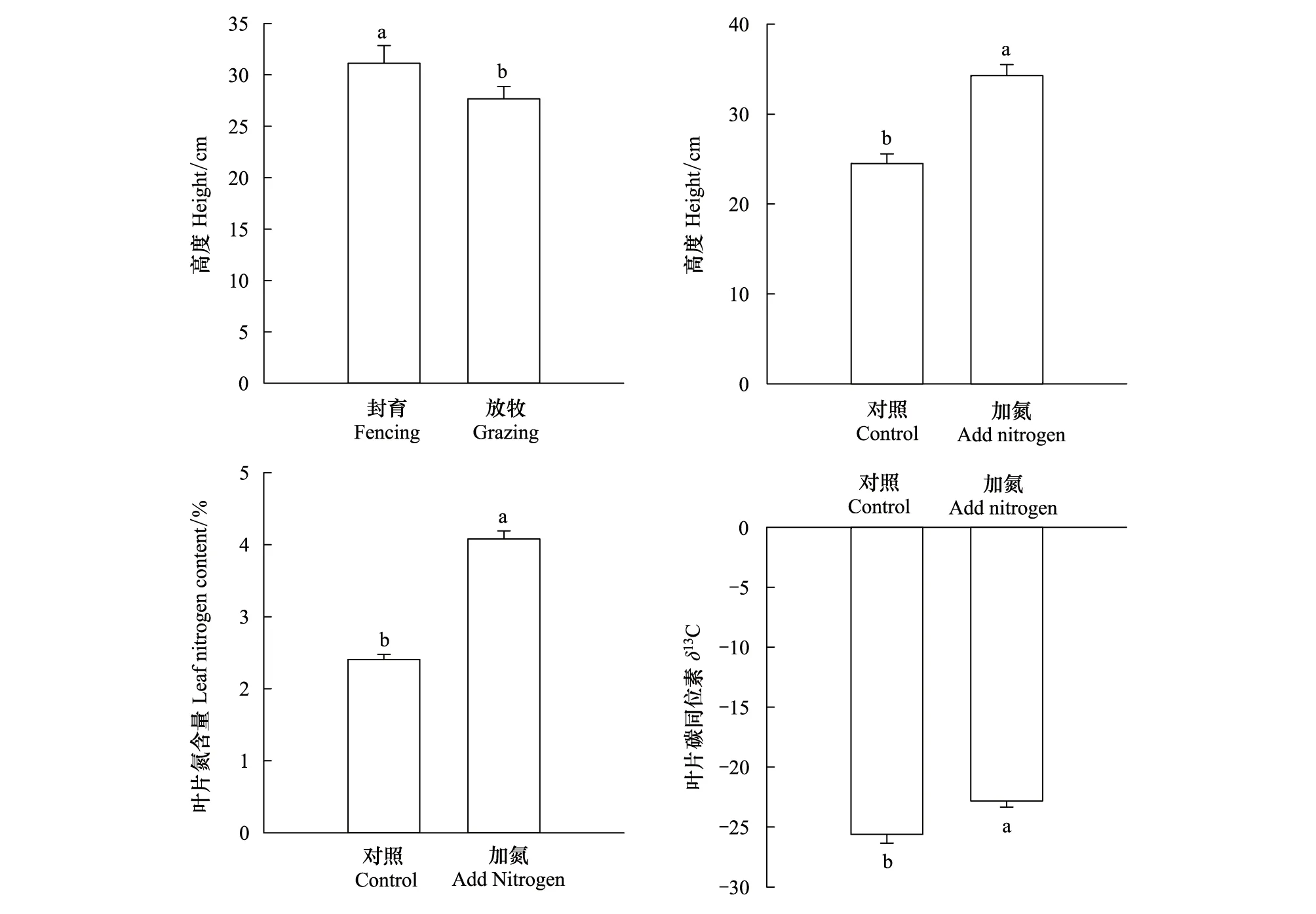
图2 放牧、氮添加对一年生植物平均功能性状的影响Fig.2 Effects of grazing and nitrogen addition on plant functional traits of annuals不同小写字母表示不同处理下一年生植物平均功能性状在P<0.05水平上的差异性
2.3 多年生植物平均功能性状对放牧、增水、氮添加及其耦合效应的响应
本研究中,氮添加对多年生植物的SLA、LNC和δ13C具有极显著影响(P<0.01),放牧和增水的耦合效应对多年生植物的SLA和LDMC具有显著影响(P<0.05),而单因素的放牧和增水对多年生植物的功能性状无显著影响(表4)。

表4 多年生植物平均功能性状对试验处理响应的方差分析Table 4 Analysis of variance (ANOVA) of perennials in responses to experimental treatments
多年生植物的SLA、LNC和δ13C在氮添加处理下分别显著升高了17.4%、32.1%和14.1%(P<0.05)。在正常降水的情况下,多年生植物的SLA在放牧处理下显著降低了14.7%(P<0.05),而多年生植物的LDMC在放牧处理下显著升高了16.4%(P<0.05),在增水情况下,放牧对多年生植物的SLA和LDMC无显著影响。在围封情况下,增水对多年生植物的SLA和LDMC无显著影响,在放牧情况下,增水对多年生植物的SLA无显著影响,而多年生植物的LDMC在增水处理下显著降低了18.4%(P<0.01)(图3)。
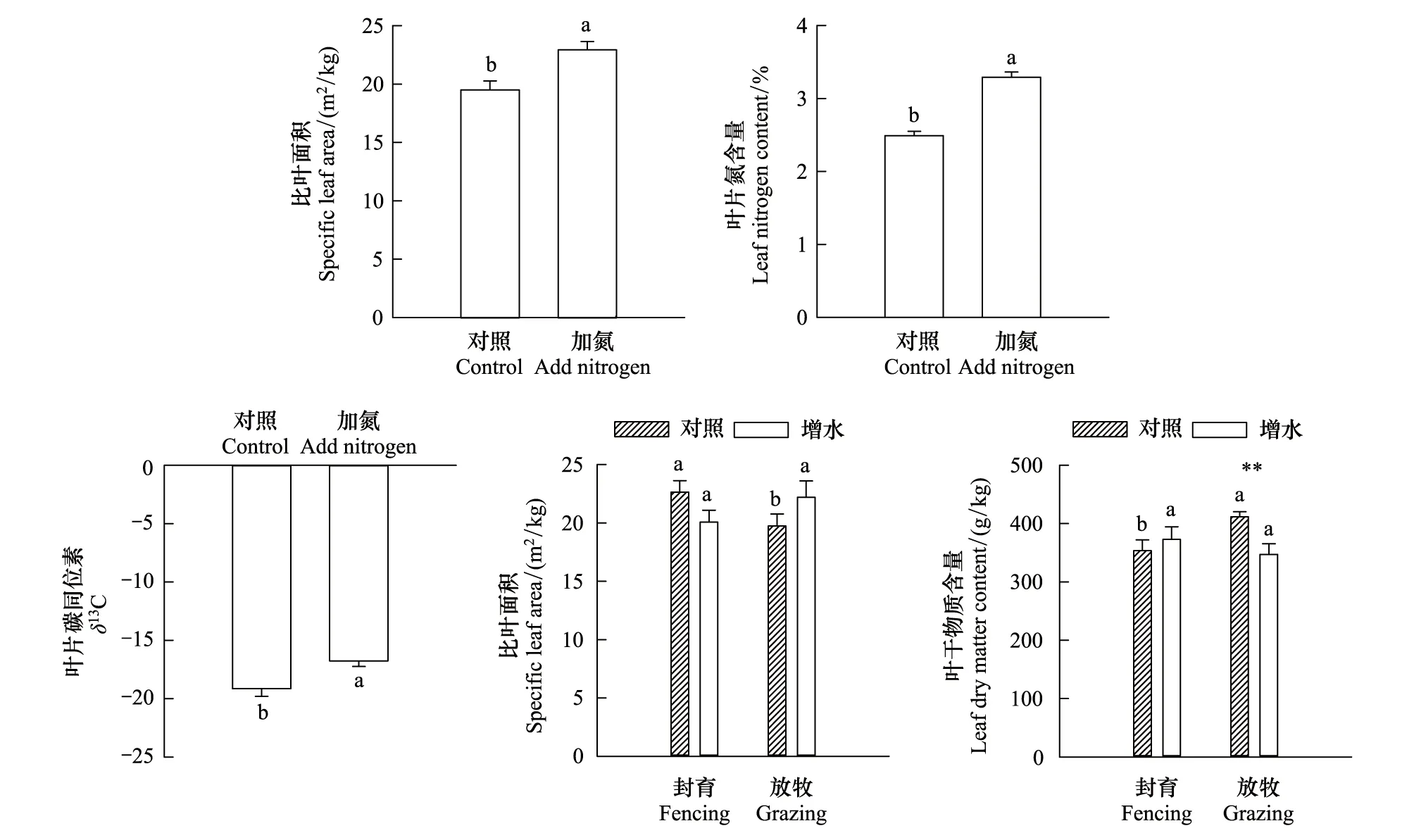
图3 氮添加、放牧和增水耦合效应对多年生植物平均功能性状的影响Fig.3 Effects of nitrogen addition, coupling effects of grazing and increase water on plant functional traits of perennials不同小写字母表示不同处理下多年生植物平均功能性状在P<0.05水平上的差异性; ** 表示放牧管理条件下增水处理对多年生植物平均功能性状在P<0.01水平上的差异性
2.4 植物群落功能性状在不同处理下的主成分分析
本研究对不同处理下沙质草地植物群落5个功能性状进行了主成分分析,结果表明:在对照处理下,CWMheight、CWMLDMC、CWMLNC和CWMδ13C在第一主成分上有较高的荷载,而CWMSLA在第二主成分上有较高的荷载,其中第一主成分的贡献率为56.4%,第二主成分的贡献率为27.8%,累计贡献率为84.3%;在放牧处理下,CWMSLA、CWMLDMC、CWMLNC和CWMδ13C在第一主成分上有较高的荷载,而CWMheight在第二主成分上有较高的荷载,其中第一主成分的贡献率为70.5%,第二主成分的贡献率为21.3%,累计贡献率为91.7%;在增水处理下,CWMLDMC、CWMLNC和CWMδ13C在第一主成分上有较高的荷载,而CWMheight和CWMSLA在第二主成分上有较高的荷载,其中第一主成分的贡献率为58.1%,第二主成分的贡献率为31.2%,累计贡献率为89.3%;在加氮处理下,CWMSLA、CWMLDMC和CWMδ13C在第一主成分上有较高的荷载,而CWMheight和CWMLNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为53.0%,第二主成分的贡献率为30.6%,累计贡献率为83.7%;在放牧和增水耦合处理下,CWMSLA和CWMδ13C在第一主成分上有较高的荷载,而CWMheight、CWMLDMC和CWMLNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为61.1%,第二主成分的贡献率为22.0%,累计贡献率为83.1%;在放牧和加氮耦合处理下,CWMSLA和CWMδ13C在第一主成分上有较高的荷载,而CWMheight、CWMLDMC和CWMLNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为61.1%,第二主成分的贡献率为30.6%,累计贡献率为91.4%;在增水和加氮耦合处理下,CWMheight和CWMLDMC在第一主成分上有较高的荷载,而CWMSLA、CWMLNC和CWMδ13C在第二主成分上有较高的荷载,其中第一主成分的贡献率为41.7%,第二主成分的贡献率为29.6%,累计贡献率为71.3%;在放牧、增水和加氮耦合处理下,CWMSLA、CWMLDMC、CWMLNC和CWMδ13C在第一主成分上有较高的荷载,而CWMheight在第二主成分上有较高的荷载,其中第一主成分的贡献率为56.2%,第二主成分的贡献率为28.1%,累计贡献率为84.3%(图4)。
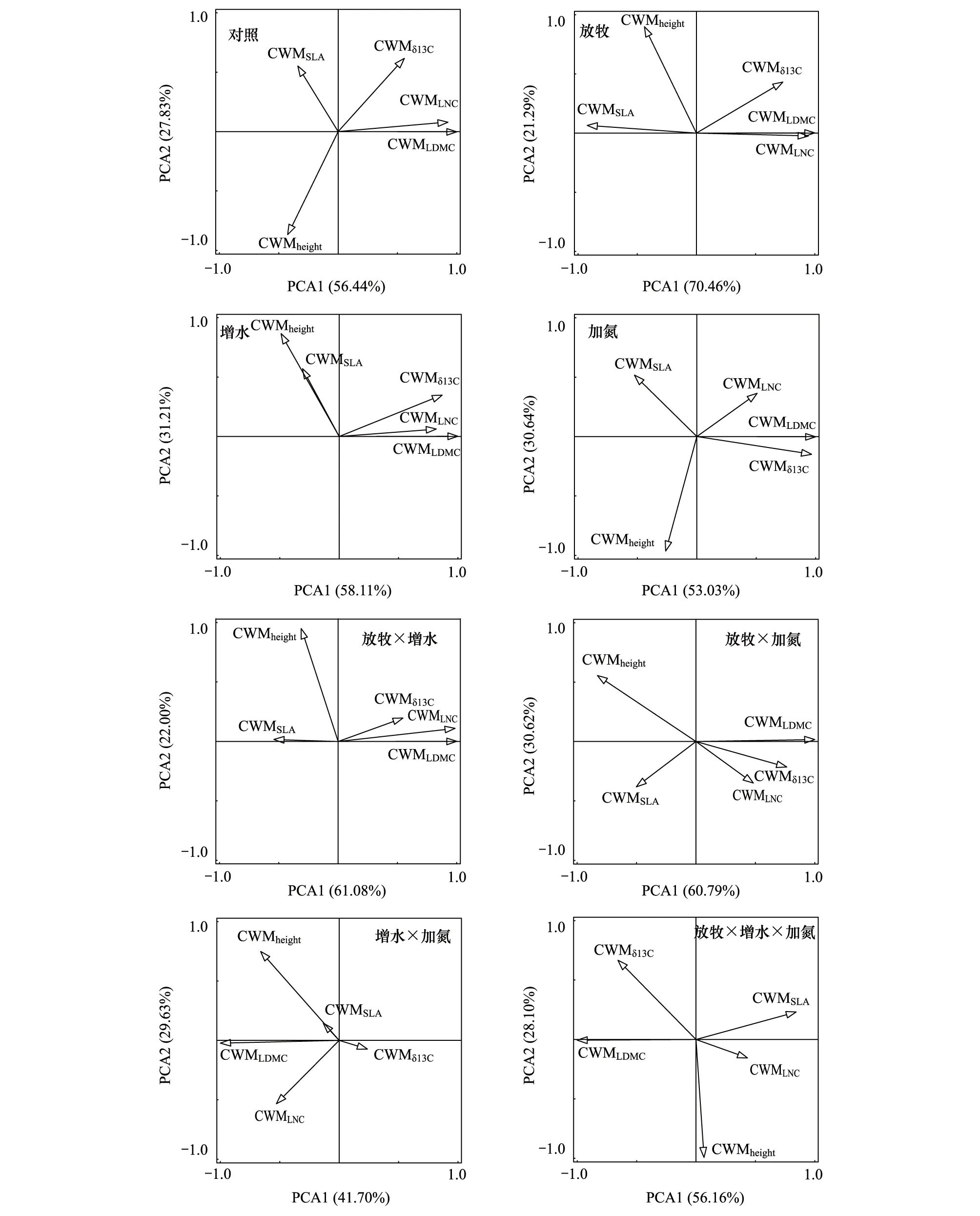
图4 植物群落功能性状在不同处理下的主成分分析 Fig.4 Principal component analysis (PCA) of plant community functional traits (CWM) under different treatmentsSLA:一年生植物的平均比叶面积 the average specific leaf area of annuals;LDMC:一年生植物的平均叶干物质含量 the average leaf dry matter content of annuals;LNC:一年生植物的平均叶片氮含量 the average leaf nitrogen content of annuals;δ13C:一年生植物的平均碳同位素the average leaf carbon isotopes of annuals
2.5 一年生植物平均功能性状在不同处理下的主成分分析
本研究对不同处理下一年生植物5个平均功能性状进行了主成分分析,结果表明:在对照处理下,植物高度、LDMC和δ13C在第一主成分上有较高的荷载,而SLA和LNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为51.5%,第二主成分的贡献率为28.9%,累计贡献率为80.4%;在放牧处理下,SLA、LDMC、LNC和δ13C在第一主成分上有较高的荷载,而植物高度在第二主成分上有较高的荷载,其中第一主成分的贡献率为74.1%,第二主成分的贡献率为21.0%,累计贡献率为95.1%;在增水处理下,LDMC、LNC和δ13C在第一主成分上有较高的荷载,而植物高度和SLA在第二主成分上有较高的荷载,其中第一主成分的贡献率为57.0%,第二主成分的贡献率为34.6%,累计贡献率为91.6%;在加氮处理下,SLA和δ13C在第一主成分上有较高的荷载,而植物高度、LDMC和LNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为51.9%,第二主成分的贡献率为35.8%,累计贡献率为87.7%;在放牧和增水耦合处理下,所有性状在在第一主成分上已经有了较高的荷载,主成分的贡献率为67.1%;在放牧和加氮耦合处理下,植物高度和LNC在第一主成分上有较高的荷载,而SLA和LDMC在第二主成分上有较高的荷载,其中第一主成分的贡献率为57.4%,第二主成分的贡献率为24.5%,累计贡献率为81.9%;在增水和加氮耦合处理下,植物高度、LNC和δ13C在第一主成分上有较高的荷载,而SLA和LDMC在第二主成分上有较高的荷载,其中第一主成分的贡献率为48.6%,第二主成分的贡献率为33.6%,累计贡献率为82.2%;在放牧、增水和加氮耦合处理下,所有性状在在第一主成分上已经有了较高的荷载,主成分的贡献率为73.3%(图5)。
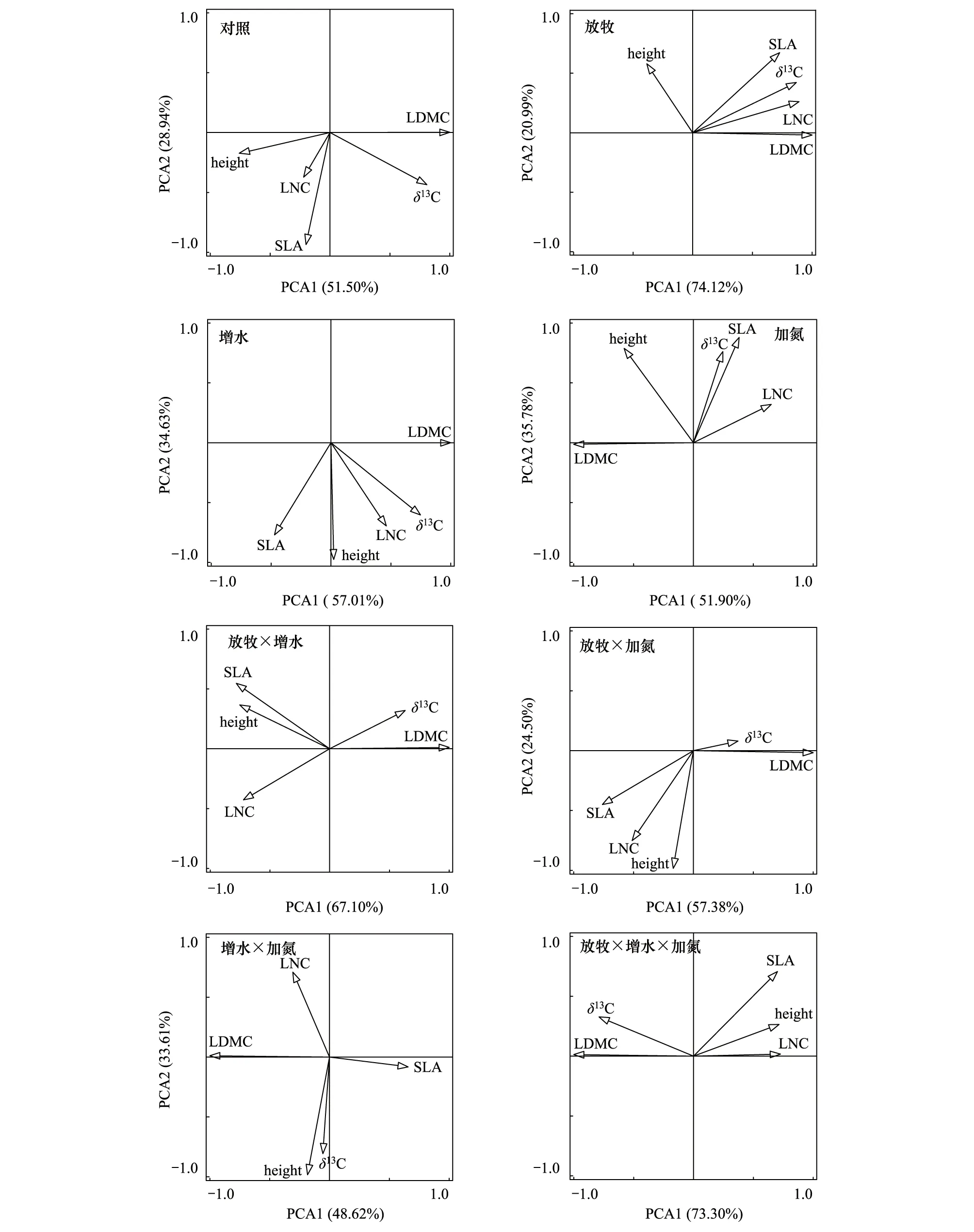
图5 一年生植物平均功能性状在不同处理下的主成分分析 (PCA)Fig.5 Principal component analysis (PCA) of average functional traits of annuals under different treatments
2.6 多年生植物平均功能性状在不同处理下的主成分分析
本研究对不同处理下多年生植物5个平均功能性状进行了主成分分析,结果表明:在对照处理下,SLA、LNC和δ13C在第一主成分上有较高的荷载,而植物高度和LDMC在第二主成分上有较高的荷载,其中第一主成分的贡献率为56.0%,第二主成分的贡献率为33.8%,累计贡献率为89.9%;在放牧处理下,植物高度、LDMC、LNC和δ13C在第一主成分上有较高的荷载,而SLA在第二主成分上有较高的荷载,其中第一主成分的贡献率为58.8%,第二主成分的贡献率为28.5%,累计贡献率为87.3%;在增水处理下,植物高度、SLA、LDMC、LNC和δ13C在第一主成分上有较高的荷载,其中第一主成分的贡献率为55.7%,第二主成分的贡献率为22.5%,累计贡献率为78.2%;在加氮处理下,植物高度、SLA和LDMC在第一主成分上有较高的荷载,而和LNC和δ13C在第二主成分上有较高的荷载,其中第一主成分的贡献率为61.3%,第二主成分的贡献率为31.8%,累计贡献率为93.1%;在放牧和增水耦合处理下,所有性状在在第一主成分上已经有了较高的荷载,主成分的贡献率为89.4%;在放牧和加氮耦合处理下,LNC和δ13C在第一主成分上有较高的荷载,而植物高度、SLA和LDMC和在第二主成分上有较高的荷载,其中第一主成分的贡献率为44.0%,第二主成分的贡献率为27.4%,累计贡献率为71.4%;在增水和加氮耦合处理下,所有性状在在第一主成分上已经有了较高的荷载,主成分的贡献率为81.8%;在放牧、增水和加氮耦合处理下,SLA、LDMC和δ13C在第一主成分上有较高的荷载,而植物高度和LNC在第二主成分上有较高的荷载,其中第一主成分的贡献率为54.5%,第二主成分的贡献率为28.9%,累计贡献率为83.4%(图6)。

图6 多年生植物平均功能性状在不同处理下的主成分分析Fig.6 Principal component analysis (PCA) of average functional traits of perennials under different treatments
3 讨论
3.1 放牧、增水、氮添加及其耦合效应对沙质草地植物功能性状的影响
外界环境变化下,不同植物通过在进化与演替过程中不断权衡和分配资源,最终形成特有的功能性状组合。研究表明,植物在放牧压力下会显现出植株变矮、节间缩短等表型性状的集合,是植物防御家畜采食、避免机械损伤的主动逃避策略[23]。本研究中,放牧对沙质草地植物群落和多年生植物的植物高度无显著影响,而显著降低了一年生植物的植物高度,这主要是由于在本试验中,放牧采取的方式为非生长季(9月到第二年3月)放牧,生长季(4月到8月)禁牧,多年生植物根系深且发达,粗根和细根分布复杂且根基深厚,非生长季遭受啃食的多年生植物在生长季禁牧的条件下容易发生补偿性生长[24],从而使草地植物群落高度维持在稳定的状态。相比于多年生植物,一年生植物多为细根且分布较浅,在放牧压力下,它们进一步丧失了阳光和土壤养分的竞争力,导致一年生植物物种组成和结构发生变化,更矮物种的种类和个数整体增加。综上,放牧降低了一年生植物的平均高度,并没有改变沙质草地植物群落的高度,而植物群落高度是影响草地生产力最主要的影响因素[25],因此,在本区域推行非生长季放牧、生长季禁牧的放牧措施能够在草地利用和维持方面找到平衡点,有利于沙质草地的可持续发展和管理。
水分是植物生长发育所必需的因子,与其他因子相比,水分对植物功能性状的影响更为强烈,如缺水会提高植物叶片的氮含量[26],常绿树种的叶片渗透水含量、叶绿素含量均会随着干季到湿季的变化而降低,而比叶重、叶片密度会随之增大[27]。然而在本研究中,增水对沙质草地植物的功能性状无显著影响,与以往研究结果存在差异的主要原因是不同地区的环境因子(温度、湿度、海拔等)具有较大差异,沙质草地中的砂土颗粒较大,土壤饱和导水率和蒸发率较高,降雨事件后土壤含水量因其持水能力较低而下降较快[28],因此,增水处理不会对沙质草地植物的功能性状产生显著影响。本研究中在正常降水下,放牧显著降低了沙质草地植物群落和多年生植物的SLA,这说明,放牧导致植物叶片具有低的SLA,从而使植物具有低的光合能力和养分周转速率,表现出植物对资源利用的“资源保持型”策略[29],而在增水30%的情况下,放牧对SLA的降低作用完全消除,这与澳大利亚东南部多年生植物的SLA随着降水的增多而升高的研究结果相一致[30],同时也强调了放牧和增水之间的耦合效应,表明增水能够减缓放牧压力对于草地植物SLA的影响。
内蒙古克氏针茅草地的研究表明,在施肥量少的处理中,克氏针茅StipakryloviiRoshev、冷蒿ArtemisiafrigidaWilld等植物的SLA和LNC相对较低,而LDMC较高,主要是养分稀缺的情况下,植物种对单位叶面积上干物质投入较高,单位叶片的光合能力和单位时间内的回报率较低;随着施肥量不断提高,植物种在单位叶面积上的干物质投入下降,叶片氮含量、单位叶片光合能力和单位时间内的回报率也逐步上升[31]。与以上研究结果相一致,本研究中氮添加显著降低了沙质草地植物群落的LDMC,说明随着氮素水平的增加,沙质草地植物群落通过改变单位面积上干物质投入来调节自身对资源的竞争[32]。此外,放牧和增水对沙质草地多年生植物的LDMC存在耦合效应,在正常降水情况下,放牧显著升高了多年生植物的LDMC含量,这体现了沙质草地中多年生植物的“资源保持型”策略,主要原因是多年生植物对于放牧的抵抗性较强,主要通过降低光合速率来应对干扰[33]。此外,拥有较高LDMC的叶片也同时有较高的木质素和纤维素,叶片硬且适口性较差,不利于反刍动物的消化代谢,是沙质草地多年生植物“耐牧”的策略[34]。然而,在增水30%的情况下,放牧对多年生植物LDMC的影响不复存在,说明增水能够减缓放牧压力对沙质草地多年生植物LDMC的影响。
氮是植物的基本营养元素,也是光合作用的底物来源[35]。研究表明,施用氮肥能够促进植物根系发育,从而有效扩大植物觅取水分和养分的土壤空间,同时还能增大蒸腾强度,降低叶水势,提高伤流液中氨基酸含量,最终提高植物吸取和运转土壤水分的能力[36]。还有研究人员构建了大气氮沉降对植物光合作用影响机理的模型,分析了全球尺度下氮添加对14种植物光合作用相关功能性状的影响。研究发现,氮添加显著增加了叶片的总面积,显著提高了植物叶片光合能力、气孔导度和蒸腾速率。随着实验持续时间增长和施氮总量增加,植物蒸腾速率的响应显著增强,表明水分消耗的增益作用随氮沉降的持续而增强[37-38]。本研究中,氮添加显著提高了沙质草地中一年生和多年生植物的水分利用效率,这与以往的研究结果相一致,表明氮添加有利于植物同化δ13C、提高叶片中δ13C的分配,最终提高植物叶片的水分利用效率。
3.2 沙质草地植物群落功能性状相关关系
有关于植物功能性状间关系的相关研究已经有很多报道。综合不同生态系统植物叶片功能性状之间的关系,总体得出叶片光合能力与LNC和SLA显著正相关[39],但是由于不同研究区域植物叶片性状之间存在着一定的差异,有些地区植物的SLA和LNC之间无相关性或呈负相关[40]。通常而言,SLA与LDMC之间为显著的负相关关系[41],主要是因为LDMC越大的叶片组织密度越高,光和CO2在叶组织中的传导受到限制[42],导致光合速率降低的同时SLA变小。植物能否适应极限环境条件,最主要的看它们能否很好地协调碳同化和水分耗散之间的关系,即植物水分利用效率是其生存的关键因子[43]。本研究通过对不同处理下沙质草地植物的不同功能性状进行主成分分析,结果表明在对照条件下,δ13C与植物高度显著负相关,其原因为具有较高δ13C的植物使用较保守的水分利用策略以允许植物生存和维持,而具有较低δ13C和挥霍的水分利用模式的植物能获得较高的生产力[44]。植物高度是直接影响草地生产力的主要因子,因此植物高度越高,水分利用效率越低,同时也表明沙质草地植物群落能够通过改变其水分利用效率适应不同的土壤水分状况,使其在植物群落中表现出更强的竞争能力和生态适应性;然而在放牧处理下,植物群落和多年生植物SLA与LDMC、LNC和δ13C之间产生了显著的负相关关系,这一结果也进一步证实SLA和LDMC间的负相关关系是陆地生态系统植物在获取养分、水分过程中不断权衡的规律。总体而言,植物群落功能性状相关性间的变化和多年生植物平均功能性状间相关性变化在不同处理下具有较为一致性的变化,说明多年生植物在调控草地植物群落应对干扰和环境变化时具有较强的主导性作用。
4 结论
放牧、增水、氮添加及其耦合效应对科尔沁沙质草地植物群落、一年生和多年生植物的功能性状产生了不同影响,具体表现为:氮添加显著提高了CWMheight和CWMLNC,降低了CWMLDMC,同时放牧和增水的耦合效应对CWMSLA具有显著影响;放牧显著降低了一年生植物的平均高度,氮添加显著提高了一年生植物的平均高度、LNC和δ13C;氮添加显著提高了多年生植物的平均SLA、LNC和δ13C,同时放牧和增水的耦合效应对多年生植物的平均SLA和LDMC具有显著影响。主成分分析表明,放牧促进了植物群落和多年生植物SLA与LDMC、LNC和δ13C的负相关关系,增水促进了植物群落和多年生植物高度与SLA的正相关关系以及LDMC、LNC和δ13C间的正相关关系,植物群落和多年生植物功能性状的相关性变化在不同处理下具有较为一致性的变化,表明多年生植物在调控草地植物群落应对干扰时具有较强的主导性作用。植物通过性状权衡将获取的资源转变为生物量,进而影响生产力和生态系统功能,然而放牧对沙质草地植物群落各个功能性状无显著影响,说明非生长季放牧、生长季禁牧有利于沙质草地的可持续发展和管理,增水能够减缓放牧压力对于草地植物的影响,氮添加有利于植物同化δ13C,并提高植物叶片的水分利用效率。
致谢:中国科学院西北生态环境资源研究院奈曼沙漠化研究站提供外业调查支持,特此致谢。
猜你喜欢
幼儿100(2020年31期)2020-11-18 03:42:00
军事运筹与系统工程(2020年2期)2020-11-16 01:11:04
文萃报·周五版(2020年29期)2020-10-20 07:11:56
疯狂英语·初中版(2019年4期)2019-09-10 07:22:44
小太阳画报(2018年6期)2018-05-14 17:19:28
军事运筹与系统工程(2018年3期)2018-03-26 06:33:02
幼儿智力世界(2017年3期)2017-04-26 23:39:37
中亚信息(2016年10期)2016-02-13 02:32:45
声学技术(2014年2期)2014-06-21 06:59:02
水土保持通报(2014年5期)2014-06-09 08:26:44
