
黄土丘陵区5种主要乔灌树种光合生理生态特征
2021-10-11 05:14:46乔雨宁任婧宇韦思瀚陈云明
水土保持研究 2021年6期
乔雨宁,任婧宇,韦思瀚,陈云明,吴 睿
(1.西北农林科技大学 水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100; 2.水利部 黄河水利委员会 黄河上中游管理局,西安 710021;3.西北农林科技大学 林学院,陕西 杨凌 712100; 4.中国科学院 水利部 水土保持研究所,陕西 杨凌 712100;5.陕西省安康市汉滨区水利局,陕西 安康 725000)
光合作用是通过光辐射作用在植物体内进行物质转化的过程,蒸腾作用是运送植物水分的主要途径[1]。光合蒸腾与植物生理因素、外界环境条件有较强的相关性,陆地植物主要以光合、呼吸作用完成生物量与生产力的累积[2]。中国学者Li[3]最早对植物光合生理特性进行了研究,表明我国3种主要气候带针阔混交林群体水平的光合特性有所差异,经过后人的持续探究,植物光合生理变化特征在乔木、灌木、草地生态群落内趋向完善[4-6]。乔木层结构复杂且丰富,灌木层是构成暖温带森林生态系统的重要部分,二者均可准确估算林下生物量,也是揭示物质能量循环机理的重要环节[7-8]。叶片作为植物与外界大气的水汽交换作用的主要器官,通过气孔开张机制将约95%水分蒸腾并散失回大气[9]。因此,研究植物叶片光合生理特性为乔灌层小型生态系统水资源配置优化、环境条件适应及能量流动循环机理提供了理论支撑。
有学者指出高温能削弱叶片光化学反应效率,阻滞CO2在叶肉细胞的交换作用[10]。另有学者基于全球尺度647套通量数据,得出植被类型对植物光合呼吸的温敏性影响最为显著[11]。我国亚热带及温带植物的在降水条件下,树木幼苗光合蒸腾速率均上升1.3倍[12-13]。当空气CO2含量倍增时,紫花苜蓿净光合速率提高17.8%,蒸腾速率降低2.5%,水分利用效率增长31%[14]。有学者研究了西北半干旱地区灌层植被梭梭、白刺及紫穗槐的光合蒸腾特性日、月变化,得出指标日变化呈“双峰”型,月变化差异性较大的结论[15]。综上所述,在不同环境条件影响下植物光合生理变化特征已有较多研究成果,但针对黄土丘陵区乡土优势乔灌树种光合反应一致性、树种搭配合理性的研究还有所不足。
自“退耕还林”工程实施以来,黄土丘陵区大面积营造人工树种,而在近20 a植被恢复的过程中,该区树种单一、搭配不合理及植被退化问题逐渐凸显,量化该区主要乔灌树种的光合生理特性能改善黄土丘陵区植被的配置空间布局。油松(Pinustabuliformis)、侧柏(Platycladusorientalis)、连翘(Forsythiasuspensa)、丁香(Syzygiumaromaticum)、紫穗槐(Amorphafruticosa)等植物作为本区优势树种,可显著提升地力,维持树种稳定[16]。目前,油松、侧柏光合蒸腾特性已有诸多研究[17],连翘、丁香、紫穗槐作为具有良好水土保持效益的灌层种,已有学者在自然立地条件下论述了其光响应过程及生理参数的变化情况[18-20],然而,目前没有将5种乔灌树种光合生理特征结合阐述的研究,故提出以下问题:(1)由于树种的不同,其趋势规律相似,但在同时段可能存在显著性差异;(2)树种光合生理特征与植物生理、气象、水分因子关联程度的高低。因此,本文以黄土丘陵区5种主要造林乔灌树种为研究对象,分析其叶片光合生理生态特征及其与生物、非生物因子的关系,旨在揭示乔灌树种光合生理特征应对外界条件变化的机制,为黄土丘陵区植被的良性恢复提供依据。
1 研究区概况
试验区位于陕西延安市安塞区纸坊沟内,位置位于109°15.17′E,36°44.90′N,海拔1 011~1 430 m,流域覆盖面积8.27 km2,属典型的黄土丘陵沟壑区,暖温带半干旱季风气候。年均温8.8℃且年较差较大,多年平均降水483.9 mm,年内及年际间降水分配不平衡,主要集中在6—8月,占全年降水的58%,太阳辐射强度高,日照总时长2 300~2 400 h。该区土壤类型以黄绵土为主,植被类型属森林草原带,主要乔木有油松、刺槐(Robiniapseudoacacia)等,灌木主要有柠条(Caraganakorshinskii)、连翘、紫穗槐等,草本植被有铁杆蒿(Artemisiasacrorum)、益母草(Leonurusartemisia)、长芒草(Stipabungeana)等。
2 材料与方法
2.1 样地布设及试验材料
在坡向(东南106°)及坡度(10°)一致的5个标准径流小区(20 m×5 m),测定当地主要造林树种,分别为2~3 a生油松、侧柏(株行距2 m×3 m)及连翘、丁香、紫穗槐(株行距1 m×2 m)纯林,对各小区内的幼树进行每木检尺,各树种分别选取长势良好的标准待测木3株作为研究对象,共计15株。
2.2 测定项目及方法
2.2.1 光合生理生态指标 于2019年7月下旬选择2个连续典型无云晴朗天气,采用LI-6400XT光合测定系统(LI-COR,Inc.,USA)测定植株光合生理指标日变化,选择自然光源叶室,测定待测木中上部受光充足的健康叶片,测定指标有:净光合速率[Pn,μmol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、水分利用效率(WUE,μmol/mmol)、气孔导度[Gs,μmol/(m2·s)]、胞间CO2浓度(Ci,mmol/mol)、气孔限制值(Ls,%)、水气压差(VpdL,kPa)、叶片温度(Tleaf,℃),利用U30-NRC小型气象站同步测定空气温度(Tair,℃)、大气CO2浓度(Ca,μmol/mmol)、空气相对湿度(RH,%)、光合有效辐射[PAR,μmol/(m2·s)]等指标。测定过程自7:00开始,19:00结束,间隔2 h测1次,每株选取3枚(簇)发育旺盛叶片,重复测定4次,取3组稳定值,平均后进行后续分析。水分利用效率(WUE)和气孔限制值(Ls)计算公式如下:WUE=Pn/Tr,Ls=1-Ci/Ca。
2.2.2 土壤体积含水量 在5种树种径流小区中部采用ECH2O土壤水分测定系统定位定点观测径流小区内土壤体积含水量的动态变化从上至下分为5个测定深度(20,50,80,120,150 cm),监测系统每30 min自动读取数据并保存。
2.3 数据分析与处理
利用Microsoft Excel 2016进行数据整理,通过SPSS 25.0对数据进行统计分析,绘图软件采用Origin 2017。通过单因素方差(One-Way ANOVA)比较5种树种光合生理指标在相同光强下的差异性,LSD法和Tamhane′s T2法对齐性、非齐性方差进行多重比较,显著性水平为p=0.05。通过Pearson法分析光合指标与生理生态因子之间的关系。通过构建回归方程揭示土壤体积含水量与光合指标之间的相关性。
3 结果与分析
3.1 光合生理参数日变化特征
连翘和丁香叶片净光合速率(Pn)日变化呈“双峰型”曲线,其中,丁香在10:00,14:00光合作用最强,Pn最大为12.61 μmol/(m2·s),连翘光合作用在10:00,14:00较强并达到峰值[10.56 μmol/(m2·s)];油松与紫穗槐日变化趋势较为相似,均为单峰曲线,其中,油松的Pn在下午14:00升至最高[3.81 μmol/(m2·s)],紫穗槐以正午12:00左右出现峰值[13.02 μmol/(m2·s)];侧柏Pn在7:00—19:00表现出持续递减趋势(图1A)。
侧柏叶片蒸腾速率(Tr)日变化呈双峰曲线,蒸腾作用在10:00及14:00左右具有较高水平,峰值为1.72 mmol/(m2·s);其他树种叶片Tr变化规律基本保持一致,7:00—14:00间Tr持续上升,且在14:00左右Tr值达到最高,随后蒸腾作用逐渐减弱,在18:00左右回落至低谷(图1B)。
侧柏、丁香、紫穗槐叶片水分利用效率(WUE)在7:00—19:00呈现递减趋势,连翘叶片水分利用状况呈“双峰”趋势,在8:00,14:00出现峰值,午间水分利用情况较差;油松叶片WUE表现出先降后升的规律,正午13:00处于水分利用较低状态;各树种叶片WUE均在7:00—9:00具有较高态势,此阶段光合作用逐步增强,气孔张度大,之后随着光照强度的增大,大气温度的上升导致叶片蒸腾作用较强,造成水分亏损,水分效率持续降低(图1C)。
油松、丁香叶片气孔导度(Gs)呈现双峰曲线,连翘、紫穗槐叶片Gs呈现单峰曲线,侧柏在7:00—19:00时Gs缓慢减小,其中,油松、丁香双峰均出现在8:00,14:00,油松、丁香Gs最大值分别为0.073,0.177 μmol/(m2·s),连翘与紫穗槐的最高值出现在11:00左右,Gs值分别为0.138,0.187 μmol/(m2·s),侧柏Gs变化范围为0.018~0.046 μmol/(m2·s)(图1D)。
侧柏、连翘及紫穗槐叶的胞间CO2浓度(Ci)值在7:00—19:00变化趋势具有一致性,均为先降后升,在14:00左右跌至谷值,油松呈现双峰曲线,双峰出现在8:00,14:00,最高峰值为331.50 mmol/mol,丁香叶片Ci变化呈“W”型曲线(图1E);3种灌木叶片在正午12:00左右出现“午休”状态,气孔限制值(Ls)均呈“双峰”型,峰值均出现在9:00,13:00,油松叶的Ls值在7:00—11:00呈迅速上升状态,午间时分降低至谷值后,14:00—18:00平缓升高,侧柏叶片Ls值在7:00—13:00呈现出先降后升的趋势且变化幅度较小,随后迅速降低至谷值(图1F)。

图1 光合生理参数的日变化曲线
3.2 光合生理参数日累积量的差异性
紫穗槐净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)日累积量均大于其余树种,说明紫穗槐有较高的生产力,其中,紫穗槐、丁香Pn,Tr累积量显著大于其余树种(p<0.05),乔木间Tr累积量差异不显著,WUE累积量仅表现出侧柏显著低于丁香、紫穗槐的规律(p<0.05)(图2A—C);丁香气孔导度(Gs)累积量在5种树种中呈现出最大值,其与紫穗槐Gs无显著性差异,而显著大于连翘、侧柏及油松的Gs累积量(p<0.05)(图2D);胞间CO2浓度(Ci)累积量表现为侧柏显著高于其余各树种的关系(p<0.05)(图2E);与Ci表现相反,仅有侧柏气孔限制值(Ls)显著低于其余树种(p<0.05),其余树种间没有显著的差异性(图2F)。
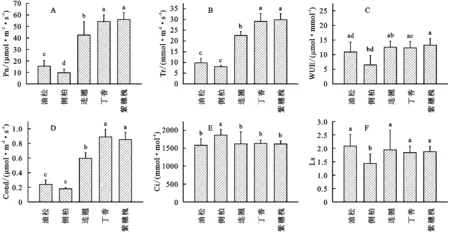
图2 光合生理参数日累积量的差异性
3.3 乔灌树种光合生理特征与影响因子的关系
3.3.1 影响乔灌树种净光合速率(Pn)的内外因素 通过Pearson相关性分析,得出了光合生理参数与生物指标、气象指标之间的关系,主要结果如下:(1)除了紫穗槐Pn与水分利用效率(WUE)间未呈现显著的相关性,其余树种两者间表现出极显著的正相关关系(p<0.01);(2)所有树种叶片Pn均与气孔导度(Gs)有极显著的相关性(p<0.01);(3)所有树种叶的Pn均与胞间CO2浓度(Ci)存在极显著负相关(p<0.01);(4)各树种叶片Pn均与气孔限制值(Ls)表现出极显著正相关;(5)侧柏Pn与叶面水汽压亏缺(VpdL)呈极显著负相关(p<0.01),而油松、连翘、丁香、紫穗槐二者间相关性不显著;(6)连翘、紫穗槐的叶片温度(Tleaf)与Pn存在一定的正相关关系(p<0.05),侧柏的二者参数间表现出极显著负相关(p<0.01);(7)空气温度(Tair)对油松、侧柏、连翘及紫穗槐叶片Pn的影响较为显著,其中,油松、紫穗槐Pn和Tair呈现显著正相关(p<0.05);连翘叶片Pn随Tair升高而升高,侧柏叶片Pn随Tair升高而降低,且呈现出极显著的相关性(p<0.01),说明侧柏最适生长温度较低;(8)大气CO2浓度(Ca)、空气相对湿度(RH)对侧柏Pn影响显著,表现为极显著正相关(p<0.01),说明Ca,RH对侧柏Pn构成间接效应,随着Ca与RH降低,引起气孔闭合,导致Pn降低,其余树种二者间相关程度较低。(9)各树种叶片Pn对光合有效辐射(PAR)变化响应显著,均呈极显著正相关(p<0.01)(表1)。

表1 各树种光合速率与影响因子的相关性
3.3.2 影响乔灌树种蒸腾速率(Tr)的内外因素 树种的Tr对植物生理因子与气象因子的响应不完全一致,植物因子中主要受到Gs和Tleaf的影响,气象因子中Tair对树种Tr影响较为明显。WUE对丁香及紫穗槐的Tr影响较大且呈极显著负相关(p<0.01);各树种Tr均与Gs有较强的联系,呈现出极显著的正相关性(p<0.01);除油松叶的Tr与Ci,Ls,VpdL未表现出相关性,其余树种在两两参数间呈现极显著的相关性(p<0.01),其中Ci与Tr呈负相关,Ls,VpdL与Tr表现出正相关性;各树种的Tr受到Tleaf的变化影响较为显著(p<0.05),均随Tleaf的升高而增大;各树种Tr与Tair呈正相关,油松、连翘、丁香及紫穗槐在二者间呈极显著相关(p<0.01);Ca,RH对油松、连翘、丁香、紫穗槐Tr的影响较为明显,均呈现出极显著的负相关性(p<0.01);仅油松Tr与PAR未表现出显著相关,其余各树种在二者参数间呈极显著正相关(p<0.01)(表2)。
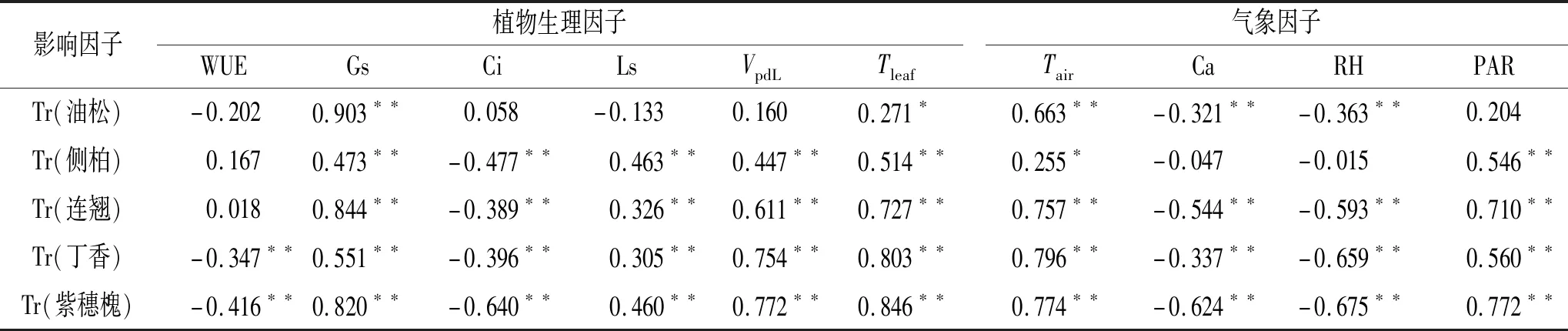
表2 各树种蒸腾速率与影响因子的相关性
3.3.3 乔灌树种净光合速率(Pn)与土壤体积含水量(SWC)的关系 通过二次曲线拟合各树种净光合速率、蒸腾速率与各层土壤体积含水量之间的关系,选取拟合结果较好(R2≥0.4)的数据组建立关系曲线,关系不显著的不展示。侧柏Pn值日变化与20,50 cm的SWC之间存在极显著的相关关系(p<0.01)(图3A—B);紫穗槐Pn值日变化与20,150 cm的SWC呈极显著相关(p<0.01)(图3C—D)。二者反映出表层土壤水分的改变易催化植物光合效能的增强与弱化。

图3 土壤水分与净光合速率之间的拟合关系
3.3.4 乔灌树种蒸腾速率(Tr)与土壤体积含水量的关系 侧柏Tr值日变化与20,50 cm的SWC之间存在极显著的相关关系(p<0.01)(图4A—B);连翘与丁香Tr值日变化分别与20,120 cm的SWC呈现出极显著的相关性(p<0.01)(图4C—D);紫穗槐Tr值日变化与20,150 cm的SWC呈极显著相关(p<0.01)(图4E—F)。

图4 土壤水分与蒸腾速率之间的拟合关系
4 讨 论
4.1 光合生理参数的日变化特征
叶片作为吸收光能及养分贮存的主要器官之一,是对外界条件改变响应最灵敏的因子[21]。紫穗槐呈单峰曲线且峰值时刻在正午,与张友焱等[15]对生长旺季紫穗槐的研究结果一致。蒋万杰等[22]发现当年与往年生油松净光合速率(Pn)日变化呈双峰曲线,与本研究油松的Pn在14:00升至最高的结果不一致,可能是由于在14:00左右的温度提高至35.904℃,光合有效辐射维持在较高水平,油松对大气温度与光照因子变化响应敏感,故此时达到峰值。侧柏Pn在7:00左右最高,原因可能是早晨CO2浓度较高,空气CO2在侧柏叶肉细胞扩散速率快,此时气孔导度值最大,气孔阻力小,进而使Pn较大,而午后Pn降至低谷,说明光强与温度都不是此时侧柏Pn的限制因素,16:00后回落至负值,说明此时侧柏呼吸速率起到主导作用,且自身开始消耗有机质维持生理作用;连翘、丁香Pn呈双峰曲线,正午时分由于过高温度、较低叶片水势、土壤水分供给不足,光合磷酸酶传递通道受阻,导致光合速率回落至低谷,蒸腾速率(Tr)是阐释植物水分损耗程度大小的关键性指标。过高蒸腾量会导致植物水分失调,对植物正常生理生长过程有限制作用,甚至达到树种的凋萎点[23]。侧柏叶片Tr呈双峰曲线,造成此现象的原因主要是正午温度高,植物由于生理作用自行关闭气孔,蒸腾作用降低。其余各树种均表现出单峰曲线且在14:00处于峰值状态,与前人结论一致[24]。正午时分,除侧柏外,其余树种在气孔作用下叶片Tr没有降低,主要是由于此时空气与叶片水汽压差较大,使得潜在蒸发速率增大,从而抵制高温破坏叶片结构。二者反映出表层土壤水分的改变易催化植物光合效能的增强与弱化。
水分利用效率(WUE)指林木损耗单位水分所统一分配的物质量,能综合反映林木光合与蒸腾能力,其与植物水资源利用度呈正比[25-26]。张慕黎等[27]发现侧柏WUE在一天内持续降低,与本研究保持一致;本研究发现丁香及紫穗槐WUE呈递减趋势,与王颖等[28]的研究结果不同,这是由于夜间的水汽聚集使得7:00空气相对湿度极大,太阳辐射强度低,蒸腾作用不明显,故此时WUE趋于峰值,之后由于温度与光强的协同效应引起植物部分气孔关闭,水分耗损持续上升,WUE趋于下降,17:00之后,光强迅速减弱,叶片对光强感应敏感,有研究表明WUE的下降与Pn的减少具有同步作用[29]。连翘、油松WUE表现出双峰曲线,与李清河等[30]研究结果一致,说明连翘作为耐旱能力较强的灌木,受到气孔因素影响较大,进而表现出“S”型曲线。
气孔导度(Gs)是测量叶片—大气水汽热平衡与碳同化的植物生理控制指标[31],油松与丁香Gs呈双峰型曲线,表明这两种树种对体内水分具有良好的调节机制,正午Gs值均回落,叶片缩小气孔张度,避免植物体水分的耗损,间接说明了油松与丁香对干旱气候的适应性较强;连翘、紫穗槐Gs值均在12:00左右达到峰值,原因可能是7月底为低矮灌木的生长旺季,水热条件充分,空气饱和水气压差较小,土壤含水量较高,表现为上午Gs持续升高,午间达到最大,之后缓慢降低;侧柏Gs值始终保持在低位水平,这与侧柏光合作用偏弱有直接联系。
Farquhar等[32]提出了气孔限制理论,他们认为胞间CO2浓度(Ci)和气孔限制值(Ls)的日变化可以分为2个阶段,午前与午后,且二者间的日变化趋势呈现出此消彼长的状态。本研究各树种Ci日变化大体呈“凹”型曲线,仅有丁香叶片Ci值在午前及午后有所波动,而Ls日变化与Ci呈现相反的变化趋势,Ci在12:00左右降至低谷可以认为气孔限制为主要原因,而午后Ci升高与Ls持续降低则表明由于叶肉细胞光合活性降低与叶绿素同化力不足而限制了光合作用,Ls在正午出现下降的趋势是由于植物的午休状态降低了气孔限制值,本研究的发现与气孔限制理论相符。
4.2 光合生理参数日累积量的差异特征
分析光合生理日累积量的差异特征能揭示特定树种在该区的生长潜力及推广应用价值。有研究表明,植物叶绿素a/b值能反映叶片对远红光的吸收能力,植物体内干物质的累积通常与供给植物生命活动能量呈反比,由于本研究两种乔木均为针叶树种,比叶面积较灌木叶片小,且灌木体内所需营养物质较乔木低,故乔木Pn显著低于灌木[33]。另有研究发现紫穗槐光合生理指标均较高[34],本研究中紫穗槐的Pn,Tr,WUE日累积量均维持在最高位水平,与前人结果一致;油松Pn日累积量显著高于侧柏,这是由于油松为弱光性树种,弱光较强光照射下生长好,侧柏为强光性树种,辐射量充足下生长较好[35],侧柏小区东西侧有山体遮蔽且内有未伐中龄侧柏导致侧柏持续接收弱光,故Pn日累积量低于油松;已有学者指出,7月连翘各项光合生理指标将达到最大值[36],而本研究中连翘Pn,Tr,Gs日累积量显著小于丁香及紫穗槐,这表明连翘光合蒸腾能力低于其余两种灌木;WUE日累积量仅表现为侧柏显著低于丁香、紫穗槐,说明侧柏生产效率及耐旱能力低于丁香、紫穗槐;本研究中3种灌木Gs累积量均显著高于乔木,说明3种灌木具有较强的气孔控制能力,有助于提高其光合机构碳同化的运转;侧柏叶片Ci日累积量显著高于其余树种,说明大气CO2在侧柏叶肉细胞扩散速率高,胞间CO2在叶绿体内扩散快,能有效抵消一定程度干旱胁迫对侧柏正常生长发育的消极影响;侧柏Ls显著小于其余树种是由于侧柏光合作用能力较弱,进入气孔的CO2较其余树种更多,水分基本能满足叶片蒸腾需求,故侧柏在进行光合作用过程中受到非气孔因素的影响更加显著。
4.3 各树种光合生理参数与影响因子的相关性
本研究中各树种Pn均值主要受到Gs,Ci,Ls、光合有效辐射(PAR)的影响,其中,Ci与各树种Pn呈现极显著负相关,其余指标表现为正相关关系,本研究5种树种叶肉细胞光合活性(RuBp羧化效率、RuBp再生速率及光合量子效率)对Pn影响较为明显,表明Gs,Ls二者均与叶片气孔起闭有密切联系。Ci与Pn变化方向相反,表明Pn会受到高CO2浓度的抑制。陈海玲等[37]认为PAR是对乔灌树种Pn起决定性作用的环境因子,与本研究保持一致。仅紫穗槐WUE与Pn关系不显著,其余树种二者间均呈极显著正相关,这表明紫穗槐水分控制能力较强,在湿润、干旱的环境均可正常生长。空气温度(Tair)、叶片温度(Tleaf)与连翘、紫穗槐Pn相关程度较高且表现出正相关性,由于温度上升能促进灌木地上受光部分的生长发育[38],连翘与紫穗槐相较其他树种具有更快的叶展开速度、更快的周转率与更强的温度调控能力[39]。油松Pn与Tair呈显著正相关,表明在生长旺季,一定范围内温度的升高能促进油松的良好生长。侧柏Pn与植物因子、气象因子均呈现出极显著的相关性,这可能由于侧柏苗木受到山体遮蔽与多年生大侧柏遮蔽的双重影响导致土壤水分偏低及弱光照射偏多故侧柏Pn受到生物、非生物因素的共同影响。
已有学者得出Tr与PAR、Tair、Tleaf、RH、土壤水势、Ci、气孔阻力、水汽压亏缺等因素有密切联系[40],而本研究中各树种Tr对外界条件的响应不完全一致,这是由于林木蒸腾特性是自身生物学特性和环境条件共同作用的结果[41]。Gs,Ci,Ls与本研究树种相关性较强,其中Ci与Tr呈负相关,这说明叶片气孔运动能显著引起林木蒸腾特性的变化,而Ci,Ls,VpdL,PAR与油松Tr未表现出显著的相关性,可能是由于研究对象为幼龄油松,针叶特性、气孔调节能力与中龄油松有所差异,具体原因有待进一步探讨。Granier等[42]认为叶片水汽压亏缺(VpdL)和PAR是短时间尺度上对蒸腾影响最关键的两个因子,这与本研究结果较为一致;RH与各树种Tr有较强负相关性,然而由于7月太阳辐射强烈,侧柏为强光性树种,通过生理调控机制,气孔关闭,RH作用气孔导度的通道受阻,此时Tr主要受气孔导度影响,故侧柏Tr与RH未表现出相关性。各树种Tr随Tleaf,Tair增大而增大,这表明在适温条件下,气孔的大幅张开加强叶片蒸腾作用,有学者认为当Tair超过林木最适光合温度后,温度过高将导致植物光合膜的热胁迫,超出有关酶的活性范围,气孔开度受到抑制,导致Tr降低[43];空气CO2浓度(Ca)与油松、连翘、丁香、紫穗槐Tr呈极显著负相关,这与叶片CO2的富集与气孔阻力大小有关。WUE仅与丁香、紫穗槐Tr呈负相关,说明丁香及紫穗槐均为喜湿灌木,植物体WUE高,故Tr低;PAR,Tleaf,VpdL直接影响气孔运动程度,进一步造成蒸腾的改变[44],故侧柏、连翘、丁香、紫穗槐PAR,Tleaf,VpdL与Tr呈极显著正相关。
Anderson等[45]研究出土壤水分(SWC)与林木光合生理生态特性有密切联系。侧柏Pn,Tr与20,50 cm的SWC呈正相关关系,说明土壤浅层水分被运输至植物体内(包括叶片),从而影响气孔运动过程及叶肉细胞中叶绿素对太阳辐射的合成,已有研究表明黄土高原侧柏的适宜生长SWC为9.78%~21.89%[46],20,50 cm的SWC在此范围内,故侧柏浅层SWC与Pn,Tr有显著的相关关系;有研究表明,当紫穗槐SWC为15%~20%,对光合蒸腾影响显著,当SWC高于20%,影响紫穗槐叶片光合蒸腾的主要因素为气象因子[47],本研究20 cm的SWC达到25%,但与Pn及Tr表现出极显著相关,这可能与7月底为生长旺季,紫穗槐生长势头好,植物体内水分循环速度较快有关;杜明新等[48]发现紫穗槐根系具有分层现象,0—40 cm以细根为主,40—80 cm以骨干根为主,150 cm土壤水势高会通过毛管力作用向骨干根转移水分,故150 cm的SWC与紫穗槐Pn及Tr相关程度较高。连翘Tr与20 cm的SWC、丁香Tr与120 cm的SWC呈极显著正相关关系,表明二者可通过调节叶片的蒸腾作用防止植株的过度失水与淹水。
5 结 论
(1)黄土丘陵区5种主要优势乔灌树种光合生理指标日变化的规律性不尽一致,其中连翘净光合速率(Pn)、水分利用效率(WUE)的日变化呈双峰曲线,紫穗槐Pn、蒸腾速率(Tr)呈“单峰”型日变化,侧柏Pn,WUE日变化表现出递减趋势,各树种气孔导度(Gs)、胞间CO2浓度(Ci)、气孔限制值(Ls)变化各异。
(2)不同树种间光合生理参数日累积量表现出一定的差异,其中,灌木Pn,Tr,WUE,Gs显著大于乔木,紫穗槐与丁香各项参数均维持较高水平,侧柏Ci显著高于其余树种,侧柏其余参数日累积量较低。
(3)5种乔灌树种平均Pn与Gs、Ci、Ls、叶片温度(Tleaf)、空气温度(Tair)、光合有效辐射(PAR)均有极显著的相关性,其中Gs与Pn的相关程度最大;平均Tr与除WUE外的其余因子相关性较强,其中Gs、叶片水汽压亏缺(VpdL)、空气相对湿度(RH)对Tr影响最为显著,Tr与Ci、空气CO2浓度(Ca)、RH呈反比;不同深度土壤含水量(SWC)对侧柏及紫穗槐影响较为显著,表层SWC(20—50 cm)与侧柏Pn,Tr呈显著正相关,150 cm的SWC与紫穗槐Pn,Tr表现出相反的趋势;浅层SWC与连翘、丁香Pn,Tr呈正比,120 cm的SWC与丁香光合蒸腾表现出极显著正相关。
本文仅简单分析了不同树种光合生理参数与影响因子的关系,未来应继续探索干旱胁迫对植株光合生理特性的影响,研究不同树种光饱和、光亏缺、弱光与强光下林木生长的快慢,细化该区环境适应性较强树种的生物生理学机理。
猜你喜欢
花卉(2024年1期)2024-01-16 11:33:50
林业科学研究(2023年4期)2023-04-29 00:44:03
新农民(2022年22期)2022-11-07 23:38:06
环境保护与循环经济(2021年5期)2021-07-23 10:16:20
山西林业(2021年2期)2021-07-21 07:29:32
星星·诗歌原创(2021年3期)2021-06-01 09:13:23
现代园艺(2017年22期)2018-01-19 05:06:56
中国饲料(2016年14期)2016-02-03 01:45:28
山西农经(2015年7期)2015-07-10 15:00:57
防护林科技(2015年3期)2015-02-22 06:18:25
