
重庆铁山坪马尾松林凋落物输入及分解动态与持水特性
2021-10-11 07:33王轶浩周建岗王彦辉
水土保持研究 2021年6期
王轶浩,周建岗,王彦辉
(1.重庆师范大学 地理与旅游学院 重庆市三峡库区地表生态过程野外科学观测研究站,重庆 401331;2.沁县漳源林场,山西 沁县 046400; 3.中国林业科学研究院 森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室,北京 100091)
凋落物层作为森林的重要水文功能层,在截持降水、减缓地表径流和防止土壤侵蚀等方面都发挥着重要作用[1]。凋落物输入及分解直接影响着凋落物现存量多少,成为决定凋落物水文功能的重要因素。因此,研究森林凋落物输入、分解对于理解和掌握凋落物水文功能的作用机理显得非常必要,并受到林学、生态学、土壤学等领域的专家学者广泛关注[2-3]。目前国内外学者对森林凋落物输入、分解、持水特征等都做了大量研究[4-6],如张建利等[7]研究认为杜鹃林的凋落物储量及持水功能受到干扰强度明显影响;侯玲玲等[8]研究认为不同群落类型的年凋落物量差异显著,并且凋落高峰出现时间以及各组分所占比例与群落类型有关;韩雪等[9]研究认为氮沉降能使凋落物分解速率减缓,且与氮沉降类型有关;武启骞等[10]研究认为红松人工林凋落物的持水性能优于蒙古栎天然林凋落物。但是,由于影响森林凋落物的因素较多,许多研究结果缺乏可比性[11],并且以往研究大都是针对凋落物现存量及持水特性或者凋落物输入、分解、持水等单个过程及特性研究,而对森林凋落物输入、分解、持水等多过程与特性研究还不足,尤其是系统开展马尾松林凋落物输入、分解及持水特性的研究鲜有报道。
马尾松(Pinusmassoniana)作为我国南方山地丘陵区主要造林树种和先锋树种,是三峡库区防护林体系中面积最大和最重要的森林类型,在保护库区生态环境方面发挥着重要作用[12]。本文在三峡库区的重庆江北区铁山坪林场,较为系统地开展马尾松林凋落物输入、分解与持水特性研究,以期揭示三峡库区马尾松林凋落物储量及其生态水文效应,为理解和评价三峡库区马尾松林水源涵养功能提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于重庆市江北区铁山坪林场(29°38′N,106°41′E)的林中园小流域,流域面积0.274 km2,属亚热带湿润气候,多年平均降水量1 100 mm,年均气温18℃。该地属四川盆东平行岭谷地貌,海拔变化为242~584 m,坡度变化为5°~30°。土壤以砂岩上发育的山地黄壤为主,土壤质地为粉砂壤土和粉砂黏壤土,厚度50~80 cm。铁山坪林场现有森林面积1 200 hm2,主要是20世纪60年代森林被破坏后天然更新形成的马尾松次生林,森林覆盖率高达90%以上。林分分层较明显,上层以马尾松为主,伴有少量香樟(Cinnamomumcamphora)、楠木(Phoebezhennan);下木层主要有木荷(Schimasuperba)、杜英(Elaeocarpussylvestris)、毛竹(Phyllostachysheterocycla)、杉木(Cuninhamalanceolata)、毛桐(Mallotusbarbatus)、白栎(Quercusfabric)等;草本层以铁芒萁(Dicranopterislinearis)为主。
1.2 研究方法
1.2.1 样地设置 在铁山坪林场林中园小流域布设4个马尾松林典型样地,林龄均为62 a左右,规格为30 m×30 m,然后调查马尾松林典型样地的基本情况和每木检尺(表1)。
1.2.2 凋落物输入监测及其现存量调查 凋落物量输入监测采用收集网法。首先在各典型样地内随机布设10个1 m×1 m凋落物收集网。收集网用孔径为1 mm的尼龙网做成,置于距离地面50 cm高处。每月按时收集凋落物2~3次,将凋落物带回实验室后,区分枯死针叶、青黄针叶、阔叶、树皮及球果、树枝、有机碎屑(包括花朵、虫尸、粪等有机体)6类凋落物组分,然后在85℃烘箱中烘干至恒重、称重,最后计算出单位面积的总凋落物量及其组分重量。
凋落物现存量测定采用烘干法。首先在各典型样地内随机布设3个1 m×1 m凋落物样方,用钢尺测量凋落物未分解层(由新鲜凋落物组成,颜色变化不明显,结构完整,无分解痕迹)、半分解层(多数凋落物已粉碎,开始分解,颜色变黑,结构已破坏,但尚可分辨)和已分解层(凋落物彻底粉碎,无法识别原有结构)厚度[7],然后分层收集,烘干称重,计算单位面积的凋落物现存量。
1.2.3 凋落物分解测定 凋落物分解采用分解袋法。首先将自然风干的马尾松未分解凋落物放在70℃烘箱中烘干后,称取50 g凋落物装入用孔径为0.5 mm的尼龙网制成的分解袋(规格为15 cm×20 cm),共36袋。2011年2月份将分解袋放在已清除地面凋落物的2号马尾松林样地内,并确保分解袋与表层土壤紧密接触,然后每月固定时间收集3袋分解袋,直到2012年2月全部回收完毕。每次将分解袋回收后,将其表面杂物清除,同样在70℃烘箱中烘干称重,其与凋落物初始干质量的差值即凋落物累积失质量,采用公式(1)—(2)计算凋落物累积失质量率和月失质量率。
(1)
(2)
式中:D为凋落物累积失质量率(%);Di为第i个月的凋落物失质量率(%);Wi为第i月凋落物失质量(g);Wi-1为第i-1个月凋落物失质量(g);W0为凋落物初始干质量(g)。
凋落物累积分解残留率与分解时间关系采用Olson负指数衰减模型(3)拟合,并采用公式(4)和(5)计算凋落物分解半衰期和全衰期。
(3)
(4)
(5)
式中:y为凋落物分解t时间的干质量残留率(%);Gt为凋落物分解t时间的干质量(g);A为拟合参数,k为凋落物年分解系数;t为分解时间(a);t0.5和t0.95为凋落物分解50%和95%时所需时间(a)。
1.2.4 凋落物持水特性测定 采用浸水法测定凋落物持水特性。首先将烘干的凋落物按未分解、半分解、未分解+半分解混合3种处理,分别称取一定重量的凋落物装入孔径为1 mm的25 cm×30 cm尼龙袋,每种处理6个重复,共18个尼龙袋。然后放入水中浸泡,分别在浸水0.5,1,2,4,6,8,10,12,24,48 h时,取出静置直到凋落物不滴水为止,称重,所得重量与烘干重量差值即凋落物在不同时间的持水量,持水量与烘干重量比值即凋落物持水率,持水率与浸水时间比值即凋落物吸水速率。将浸水48 h的凋落物持水率视为最大持水率,并根据凋落物现存量计算凋落物最大持水量和最大持水深。凋落物有效拦蓄量采用公式(6)计算得到。
Ws=(0.85Rmax-R0)×M
(6)
式中:Ws为凋落物有效拦蓄量(t/hm2);Rmax为凋落物最大持水率(%);R0为凋落物自然含水率(%);M为凋落物现存量(t/hm2)。
2 结果与分析
2.1 凋落物输入及现存量
森林凋落物输入不仅与树种组成、林龄、密度及其生物节律等特征有关,还受到自然环境条件明显影响[13],如干旱、大风等。由表2可知,2011年马尾松林总凋落物量为493.44 g/m2,其中,以枯死针叶最多,为231.35 g/m2,占总凋落物量46.89%;其次是有机碎屑,为103.57 g/m2,占总凋落物量20.99%;以青黄针叶最少,为11.55 g/m2,占总凋落物量2.34%。马尾松林凋落物不同组分排序为枯死针叶>有机碎屑>树枝>阔叶>树皮及球果>青黄针叶。
马尾松林月总凋落物变化在11.54~90.31 g/m2,基本呈双峰模式,其中,第1高峰发生在4—5月份,总凋落物为129.13 g/m2;第2高峰发生在7—9月份,总凋落物为146.7 g/m2,两个高峰的总凋落物为275.83 g/m2,占年总凋落物的69.89%。马尾松林总凋落物第1高峰的出现,可能是因为4月、5月份正值马尾松开花期和阔叶树种换叶期。由表2可知,4月、5月份有机碎屑的月凋落物分别为22.26,22.28 g/m2,处于其年内变化的第三、二位,共占其年凋落物的43%和总凋落物第1高峰的34.49%。4月、5月份阔叶凋落物达到其月凋落物年内变化的最高峰,分别为9.55,6.77 g/m2,共占其年凋落物41.63%。马尾松林总凋落物的第2高峰出现,可能是因为马尾松林受到干旱环境胁迫影响[13],其中7月、8月份正值重庆地区的伏旱季节。7—9月份枯死针叶凋落物达到其年内变化的峰值(99.82 g/m2),占其年凋落物的42.96%和总凋落物第2高峰的36.19%。同样地,树枝凋落物在该时段达到其年内变化的峰值,其中7月、8月份树枝分别为18.05,12.28 g/m2,共占其年凋落物的40.80%。
青黄针叶为非正常凋落的针叶,更能反映针叶对自然环境胁迫的响应,本研究青黄针叶月凋落物变化在0.15~3.71 g/m2,也在7月、8月份的伏旱季节达到其年内变化的峰值,分别为3.71,1.89 g/m2,共占其年凋落物的48.48%,同样可说明马尾松林凋落物受干旱环境明显影响。
总体上,除阔叶凋落物的年内变化呈单峰模式外,其他组分的年内变化趋势与总凋落物一致,均呈双峰模式,这可能是因为马尾松林中阔叶凋落物的年内变化只受其生物节律支配,而马尾松凋落物的年内变化除受其生物节律支配外,还受到干旱等外界环境明显影响[13]
由表3可知,马尾松林凋落物现存量为1 204 g/m2,其中以已分解层凋落物最多,为463 g/m2,占凋落物现存量38.46%;其次是半分解层,为380 g/m2,占凋落物现存量31.56%;以未分解层凋落物最少,为361 g/m2,占凋落物现存量29.98%。同时,结合表2可知,马尾松林凋落物的年输入量占现存量的40.98%。

表3 马尾松林凋落物现存量
2.2 凋落物分解
凋落物分解直接决定着凋落物现存量及其层次组成与分配,从而影响着凋落物层的生态水文功能。由表4可知,马尾松林凋落物的累积失质量和累积失质量率均随分解时间延长而增大,其中,马尾松林凋落物分解1个月后,其累积失质量和累积失质量率分别为1.85 g,3.71%;分解半年后,其累积失质量和累积失质量率分别为11.36 g,22.72%,是年失质量(17.81 g)和年失质量率(35.62%)的63.78%,这可能是3—8月份的温度、湿度等自然环境条件比较适宜马尾松林凋落物分解所致。从马尾松林凋落物月失质量率也可看出(表4),3—8月份月失质量率普遍较高,除7月份为1.55%外,其余月份均在2%以上,平均为3.79%。而9—次年2月份的月失质量率普遍较低,除9月份、12月份外,其余月份基本在1.5%以下,平均为2.15%。马尾松林凋落物月失质量率年内变化为1.04%~6.75%,最大值发生在8月份,最小值发生在2月份,平均为2.97%。

表4 马尾松林凋落物分解特征
由图1可知,马尾松林凋落物干质量残留率(y)与分解时间(x)呈显著指数函数关系,随分解时间增大而减小,拟合方程为y=99.414e-0.46x(R2=0.9855,p<0.01,n=12)。分解1 a后,马尾松林凋落物干质量残留率为64.38%,进一步研究表明马尾松林凋落物年分解系数为0.46,分解50%(半衰期)为1.51 a;分解95%(全衰期)为6.51 a。
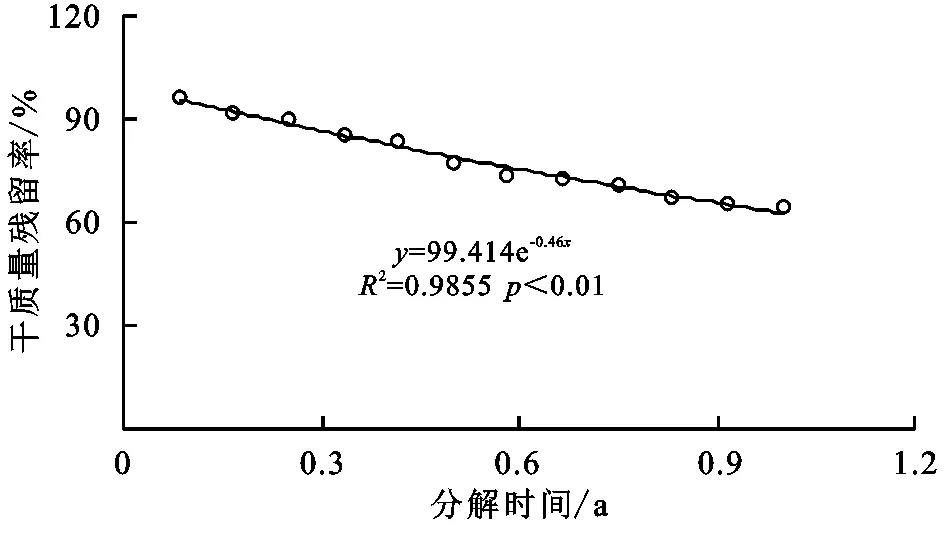
图1 马尾松林凋落物干质量残留率和分解时间的关系
2.3 凋落物持水特征
马尾松林未分解、半分解、未分解+半分解凋落物的持水率均随浸水时间增加而增大,呈倒“J”型,其持水过程可划分为快速增加阶段(0~4 h)、缓慢增加阶段(4~12 h)、趋于稳定阶段(12~48 h)3个阶段(图2)。在浸水0.5 h时,持水率即分别达到104.72%,160.61%,141.14%;在浸水4 h时,持水率则分别为139.18%,184.08%,169.63%,相比0.5 h时分别增加了34.46%,23.47%,28.49%。总体上,不同浸水时间的马尾松林凋落物持水率均表现为半分解>半分解+未分解>未分解,说明半分解凋落物的持水能力优于其他层次凋落物。由图2可知,马尾松林未分解、半分解、未分解+半分解凋落物的吸水速率随浸水时间增加而减小,呈“J”型。同样地,马尾松林凋落物吸水过程也可划分为急剧减小阶段(0~4 h)、缓慢减小阶段(4~12 h)、趋于稳定阶段(12~48 h)3个阶段。在快速减小阶段,未分解、半分解、未分解+半分解凋落物的吸水速率由0.5 h时2.10,3.15,2.82 kg/(kg·h)分别下降到4 h时0.36,0.46,0.42 kg/(kg·h),仅为0.5 h的1/7左右;在趋于稳定阶段,各层次凋落物的吸水速率最后接近于“0”。这说明马尾松林凋落物在0.5 h内发挥的水文功能最强,对短时降水的截持调蓄功能也最大,其后则急剧下降,4 h之后其截持调蓄能力趋于饱和,12 h之后则完全饱和。
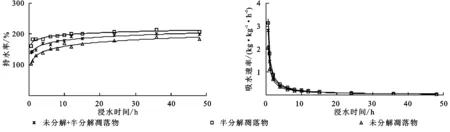
图2 马尾松林凋落物持水和吸水过程
马尾松林未分解、半分解、未分解+半分解凋落物的持水率与浸水时间均呈极显著正相关关系(p<0.01,表5),可用对数函数拟合:s=alnt+b,式中:s为持水率(%);t为浸水时间(h);a为方程系数;b为方程常数项。由表5可知,马尾松林各凋落物的吸水速率与浸水时间则均呈极显著负相关关系(p<0.01),可用幂函数拟合:v=at-b,式中:v为吸水速率[kg/(kg·h)];t为浸水时间(h);a为方程系数;b为方程常数项。

表5 马尾松林凋落物持水率、吸水速率与浸水时间的拟合方程
最大持水能力是评价凋落物水源涵养功能的最重要指标,由表6可以看出,马尾松林未分解层、半分解层、未分解+半分解层凋落物的最大持水量分别为6.63,7.84,14.64 t/hm2,最大持水深分别为0.66,0.78,1.46 mm。有效拦蓄能力则更能反映凋落物实际截持调蓄降水的效果,它与凋落物初始湿度密切相关,研究表明,马尾松林未分解层、半分解层、未分解+半分解层凋落物的有效拦蓄率分别为139.37%,154.14%,150.52%,有效拦蓄水深分别为0.50,0.59,1.12 mm。

表6 马尾松林凋落物持水特征
3 讨 论
树木凋落物多少由其本身生物学特性和自然环境条件共同影响,不同树种的凋落物量及其凋落节律各不相同[10-11],本研究马尾松针叶凋落主要发生在7—9月份,而阔叶凋落主要发生在3—5月份。森林凋落物多少则主要取决于森林类型及树种组成、起源、林龄、密度等林分特征和气候、土壤、地形等自然环境因素,不同立地条件、森林类型的凋落物量差异很大[14],但其年内月变化规律一般呈“单峰”型、“双峰”型或不规则类型[13]。本研究马尾松林年凋落物量为4.93 t/hm2,高于郭婧等[11]对中亚热带杉木人工林研究结果(4.14 t/hm2),但低于中亚热带石砾+青冈常绿阔叶林(7.24 t/hm2)、南酸枣落叶阔叶林(7.95 t/hm2)和马尾松针阔混交林(8.18 t/hm2)。此外,本研究马尾松林总凋落物年内变化呈双峰模式,而除阔叶外,其他组分凋落物年内变化规律也呈双峰模式,这与中亚热带杉木人工林和南酸枣落叶阔叶林一致[11]。
凋落物现存量由凋落物输入和分解共同决定,处于动态平衡,并直接影响着凋落物层水文功能。本研究马尾松林现存量为12.04 t/hm2,与贾秀红等[15]研究结果相近(马尾松林未分解层和半分解层凋落物储量为6.32 t/hm2),但要低于鼎湖山马尾松林(21.96 t/hm2)[6]、贵州龙里林场马尾松林(32.20 t/hm2)[16]、重庆四面山马尾松+杉木+木荷混交林(64.47 t/hm2)[17]和三峡库区马尾松林枯落物贮存量平均值(15.70 t/hm2)[18]。这可能与本研究马尾松林林分密度较小以及受酸沉降影响而针叶凋落物年输入较少[13]有关。
凋落物分解主要是由温度、湿度、酸碱度等外界环境条件以及凋落物组分通过影响土壤生物活动而起作用的,不同森林类型凋落物的分解特征不同,一般阔叶林>针阔混交林>针叶林[19]。我国针叶林凋落物年失质量率变化在20%~40%,阔叶林凋落物变化在43.5%~85%[20]。本研究马尾松林年失质量率为35.62%,半衰期为1.51 a,全衰期6.51 a,这与任来阳等[21]在该地区的研究结果相近,但高于三峡库区秭归马尾松人工林(30.4%)[3],明显低于福建洋口林场马尾松人工林(年失质量率为91.6%,半衰期为4.7个月,全衰期为19个月)[22]、江西千烟洲马尾松林(50%)[23]和广西禄峰山林场马尾松林(全衰期为2.13~2.46 a)[24],这可能是由本研究马尾松林的土壤酸化严重、土壤动物和微生物种类和数量减少所致[21,25]。
凋落物以其疏松的内部结构和巨大的表面积,在截持降水、增加降雨入渗、减少地表径流、防止土壤侵蚀等方面都具有重要作用[10]。研究表明,不同浸水时间的马尾松林凋落物持水率均以半分解最大,未分解最小,未分解+半分解居中,这与魏强等[14]研究结果一致,主要是因为相比未分解凋落物,马尾松林的半分解针叶凋落物含油率更高、表面积更大。凋落物持水能力则由凋落物现存量和持水率共同决定,本研究马尾松林未分解、半分解、未分解+半分解层凋落物的最大持水深分别为0.66,0.78,1.46 mm,均明显低于其他研究地区的马尾松林凋落物[16-18],这主要与本研究马尾松林的凋落物现存量较少及其最大持水率较小有关。
4 结 论
(1)马尾松林总凋落物量为493.44 g/m2,为现存量(1 204 g/m2)的40.98%;马尾松林月总凋落物变化在11.54~90.31 g/m2,与其他组分(除阔叶外)凋落物的年内变化趋势均呈双峰模式,年凋落物组分排序为枯死针叶>有机碎屑>树枝>阔叶>树皮及球果>青黄针叶。
(2)马尾松林凋落物年失质量率为35.62%,半衰期为1.51 a,全衰期为6.51 a;月失质量率变化在1.04%~6.75%,平均为2.97%,以3—8月份的月失质量率普遍较高;凋落物的干质量残留率与分解时间呈显著负相关(p<0.01)。
(3)马尾松林未分解层、半分解层和未分解+半分解层凋落物的最大持水量分别为6.63,7.84,14.64 t/hm2,有效拦蓄水深分别为0.50,0.59,1.12 mm;各凋落物的持水率均随浸水时间呈倒“J”型变化,不同浸水时间的凋落物持水率均为半分解>半分解+未分解>未分解,吸水速率则均随浸水时间呈“J”型变化。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
戏曲研究(2022年2期)2022-10-24
广西植物(2021年9期)2021-10-16
农家科技中旬版(2020年10期)2020-07-12
西夏研究(2020年1期)2020-04-01
农家科技中旬版(2020年2期)2020-03-18
绿色科技(2019年5期)2019-11-29
翠苑(2019年4期)2019-11-11
收藏界(2019年4期)2019-10-14
作文周刊·小学四年级版(2019年8期)2019-04-23
