
胡黄连苷对SH-SY5Y细胞氧糖剥夺/复氧复糖损伤保护作用
2021-10-09 19:05王潇璐于竹芹汪贯习刘宗保周缜郭云良
青岛大学学报(医学版) 2021年4期
王潇璐 于竹芹 汪贯习 刘宗保 周缜 郭云良
[摘要] 目的 探讨胡黄连苷对人神经母细胞瘤SH-SY5Y细胞氧糖剥夺/复氧复糖(OGD/R)损伤的保护作用及其最佳剂量。方法 体外培养SH-SY5Y细胞,建立OGD/R模型。于复氧复糖时分别加入胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ各5个浓度梯度(5、10、20、40、80 μmol/L),优化最佳剂量。后续选用胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ各自的最佳剂量,进行2×2×2析因设计分组。光学显微镜下观察各组细胞形态学变化,采用CCK-8法检测细胞存活率,采用乳酸脱氢酶(LDH)释放法测定细胞损傷程度,采用荧光探针DCFH-DA标记法检测细胞内活性氧(ROS)的水平,根据用药剂量及结果综合评价胡黄连苷的保护作用。结果 与对照组相比较,OGD/R后SH-SY5Y细胞的形态结构发生改变,模型组细胞变圆或肿胀,突起回缩或消失,细胞膜有皱褶或破损,细胞数目明显减少,漂浮细胞增多;与模型组相比较,胡黄连苷组细胞损伤得到不同程度的改善,细胞贴壁性增强。对各指标进行综合分析,从用药剂量最小化和疗效最大化的角度考虑,选用胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ的最佳剂量为20 μmol/L进行后续实验。胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ联合应用组与其他组相比,细胞形态较完整,细胞贴壁性强,细胞存活率明显提高,LDH含量下降,细胞损伤明显减轻,ROS水平下降(F=82.9~186.3,P<0.01)。结论 胡黄连苷可能通过抗氧化应激对SH-SY5Y细胞OGD/R损伤发挥神经保护作用,且胡黄连苷Ⅰ、Ⅱ、Ⅲ各20 μmol/L时联合使用效果最好。
[关键词] 胡黄连甙;再灌注损伤;卒中;神经保护;氧化性应激
[中图分类号] R743.3
[文献标志码] A
[文章编号] 2096-5532(2021)04-0537-07
脑卒中是临床上常见的脑血管疾病,其中缺血性脑卒中约占80%[1]。东亚人群患脑卒中的风险位居全球首位[2-3]。脑缺血再灌注损伤是指脑组织缺血后血流再通进一步导致组织损伤和功能障碍的过程,是一种涉及多个因素的复杂病理生理现象[4]。氧糖剥夺/复氧复糖(OGD/R)神经细胞模型,是通过氧糖剥夺后快速恢复氧糖供应,从而在细胞水平模拟临床脑缺血再灌注损伤,以进行细胞机制和药物研究[5]。正常情况下,自由基的生成和清除会形成动态平衡。在缺血再灌注时,会产生大量的自由基[6-7]。大量的活性氧(ROS)是导致缺血再灌注损伤的主要原因[8-9]。ROS是有氧代谢产生的自由基,是一种不稳定的化合物质,易与核酸、脂质和蛋白质反应,从而引起细胞损伤或细胞死亡[10]。ROS导致的氧化应激会进一步引起DNA损伤和局部炎症反应,引发炎症级联反应可能导致随后的炎症因子风暴,使细胞结构受损,最终导致细胞死亡[11-12]。因此,研究开发抗氧化应激的神经保护药物至关重要。传统中药胡黄连是玄参科植物西藏胡黄连的干燥根茎[13]。研究证明,胡黄连的主要有效成分是环烯醚萜类化合物,主要单体有效成分为胡黄连苷Ⅰ、胡黄连苷Ⅱ和胡黄连苷Ⅲ;胡黄连苷具有保护肝脏[14],保护脑缺血再灌注[15-17]、肾缺血再灌注、急性胰腺炎[18]以及减轻炎症反应[19]的作用。但胡黄连苷对SH-SY5Y细胞OGD/R损伤的保护作用和最佳剂量尚未见报道。本研究通过建立人神经母细胞瘤SH-SY5Y细胞OGD/R模型模拟脑缺血再灌注损伤过程[20-21],旨在探讨胡黄连苷对OGD/R损伤的保护作用及其最佳剂量,以期为缺血性脑血管病的治疗提供新的思路。
1 材料与方法
1.1 药品和试剂
胡黄连苷Ⅰ(分子式为C24H28O11,分子量为492.47)、胡黄连苷Ⅱ(分子式为C23H28O13,分子量为512.46)、胡黄连苷Ⅲ(分子式为C25H30O13,分子量为538.5)(B20515,上海源叶生物科技有限公司)。DMEM高糖培养基(Hyclone, USA);DMEM无糖培养基(GIBCO,USA);胰蛋白酶(Trypsin,GIBCO,USA);澳洲胎牛血清(GIBCO,USA);细胞培养专用100×青-链霉素混合液(P1400,Solarbio);Cell counting kit-8(DOJINDO,Japan);乳酸脱氢酶(LDH)检测试剂盒(南京建成生物工程研究所,中国);ROS检测试剂盒(碧云天,中国)。
1.2 SH-SY5Y细胞培养
人神经母细胞瘤SH-SY5Y细胞株(批号为20181105)由中国医学科学院基础医学研究所细胞资源中心提供,置于含有体积分数0.10胎牛血清、100 kU/L青霉素和100 g/L链霉素的DMEM培养基中培养。细胞接种后放置在37 ℃、体积分数0.05 CO2培养箱中,待生长至对数期时作为实验细胞。当细胞覆盖瓶底面积达80%时,倒掉培养液,用PBS清洗,加入2.5 g/L的胰蛋白酶,消化30 s后以1 000 r/min离心5 min,使用完全培养基重悬,将细胞悬液以2×108/L接种于96孔板中,置于37 ℃、体积分数0.05 CO2培养箱中过夜培养。
1.3 OGD/R模型制备
细胞生长至汇合率为80%时,将含有体积分数0.10胎牛血清的DMEM高糖培养基更换成无糖无血清的DMEM培养基,置于37 ℃培养箱中,连续充以无氧气体(体积分数0.90的N2、体积分数0.09的CO2和体积分数0.01的O2),4 h后取出,换成含体积分数0.10胎牛血清的DMEM高糖培养基,放入37 ℃、体积分数0.05 CO2的培养箱中培养24 h。
1.4 分组设计
前期实验将细胞随机分为对照组(使用含体积分数0.10胎牛血清的DMEM高糖培养基进行正常培养,不做其他处理)、模型组(建立OGD/R模型)、胡黄连苷组(建立OGD/R模型,在复氧的同时加入5、10、20、40、80 μmol/L等5个浓度梯度的胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ)。每次每组设5个平行复孔,实验重复3次。通过观察细胞形态和检测细胞存活率、LDH释放量和ROS水平,确定胡黄连苷Ⅰ、胡黄连苷Ⅱ和胡黄连苷Ⅲ的最佳浓度。
后续实验选用胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ的最佳浓度作为研究因素,采用2×2×2的析因设计分组,每个因素各取2个水平,即胡黄连苷Ⅰ(0用、1不用)、胡黄连苷Ⅱ(0用、1不用)、胡黄连苷Ⅲ(0用、1不用),不同水平完全交叉组合形成了23个实验组,即8个实验组。
1.5 检测指标
1.5.1 细胞生长状况 复氧复糖24 h后,在倒置显微镜下每组随机选取5个视野拍照,观察细胞的生长状况、数量、形态的变化。
1.5.2 CCK-8法检测细胞存活率 将生长状态良好的细胞消化、离心,用培养基重悬细胞、计数。取细胞悬液每孔100 μL(2×108/L)铺在96孔板中,放入37 ℃、含体积分数0.05 CO2培养箱中培养24 h,待细胞汇合率达到80%时,制备OGD/R模型。造模结束后,在每孔中加入10 μL的CCK-8溶液,放入培养箱中孵育3 h,同时再设一个空白孔作为对照,用全自动酶标仪检测450 nm处各孔的吸光度值。计算各组SH-SY5Y细胞的存活率,细胞存活率=实验组吸光度均值/对照组吸光度均值×100%。实验重复3次。
1.5.3 LDH释放量测定 SH-SY5Y细胞分组及培养同前,在OGD/R后,于96孔板中取各组处理后的细胞培养上清液适量,进行LDH测定。反应结束后用酶标仪测定490 nm处的吸光度值,计算细胞损伤程度。
1.5.4 细胞内ROS水平检测 SH-SY5Y细胞分组及培养同前,在OGD/R后,弃去细胞培养上清液,加不含血清DMEM培养基稀释的DCFH-DA工作液,37 ℃孵育30 min,弃去上清液,洗涤2次,使用PBS吹打均勻,应用荧光酶标仪进行检测,设置激发波长为488 nm、发射波长为525 nm。结果以荧光强度比值表示,荧光强度比值=实验组的荧光强度/对照组的荧光强度。
1.6 统计学处理
应用Graph Pad Prism 8.0统计学软件进行数据处理。计量资料结果以X2±s表示,胡黄连苷对OGD/R后SH-SY5Y细胞的存活率、损伤程度和ROS水平影响的分析采用单因素方差分析(One-way Anova),组间两两比较采用LSD t检验;胡黄连苷组合优化对OGD/R后SH-SY5Y细胞的影响分析采用析因设计的方差分析。以P<0.05为差异有统计学意义。
2 结 果
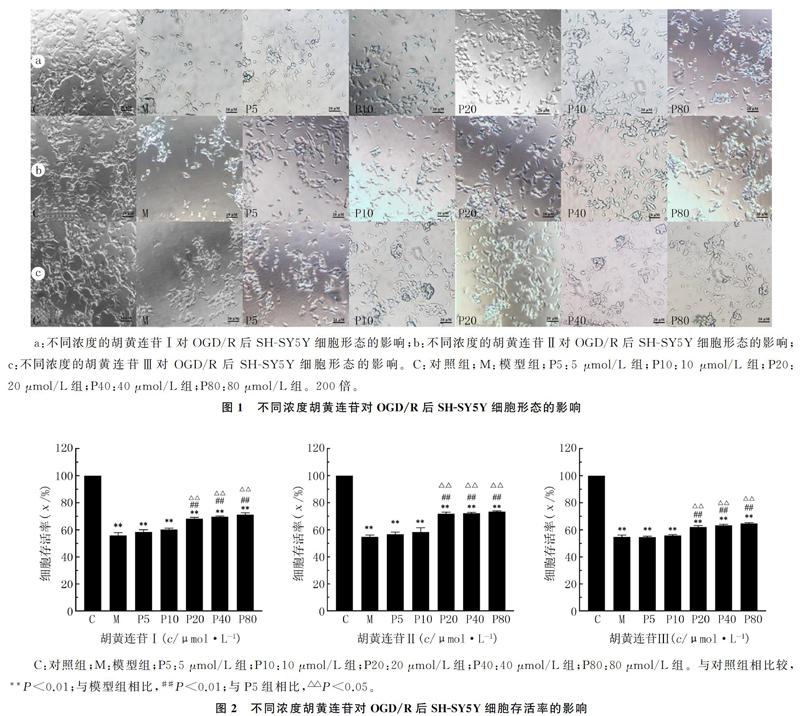
2.1 胡黄连苷对OGD/R后SH-SY5Y细胞形态学的影响
对照组细胞大部分正常贴壁生长,仅少量半贴壁,漂浮细胞数量较少,细胞形态呈两极或多极,细胞突起明显,细胞之间相互交织成网状,细胞膜光滑、完整,胞体折光性较强。与对照组相比较,模型组细胞的间隙变大,部分细胞形状不规则,细胞皱缩,细胞质凝聚,细胞突起明显减少,贴壁细胞数量明显减少,部分细胞团脱落,胞体折光性下降,部分细胞裂解成碎片。胡黄连苷组在药物浓度为20~80 μmol/L时细胞状态较好,OGD/R导致的SH-SY5Y细胞形态变化明显减轻。虽然细胞间隙增大、突起有所回缩,但是细胞数量比模型组明显增加,细胞贴壁性增强,可观察到正常形态细胞,大部分细胞呈长梭形或三角形,少部分细胞皱缩变形或脱壁,细胞碎片与模型组相比明显减少,生长状态有明显改善。其中药物浓度为20、40 μmol/L时细胞形态较好、贴壁能力较强,两个浓度组细胞形态无明显差异;当药物浓度为80 μmol/L时细胞间隙增大。根据剂量最小化和疗效最大化的原则,胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ均选择以20 μmol/L为最佳剂量。见图1。
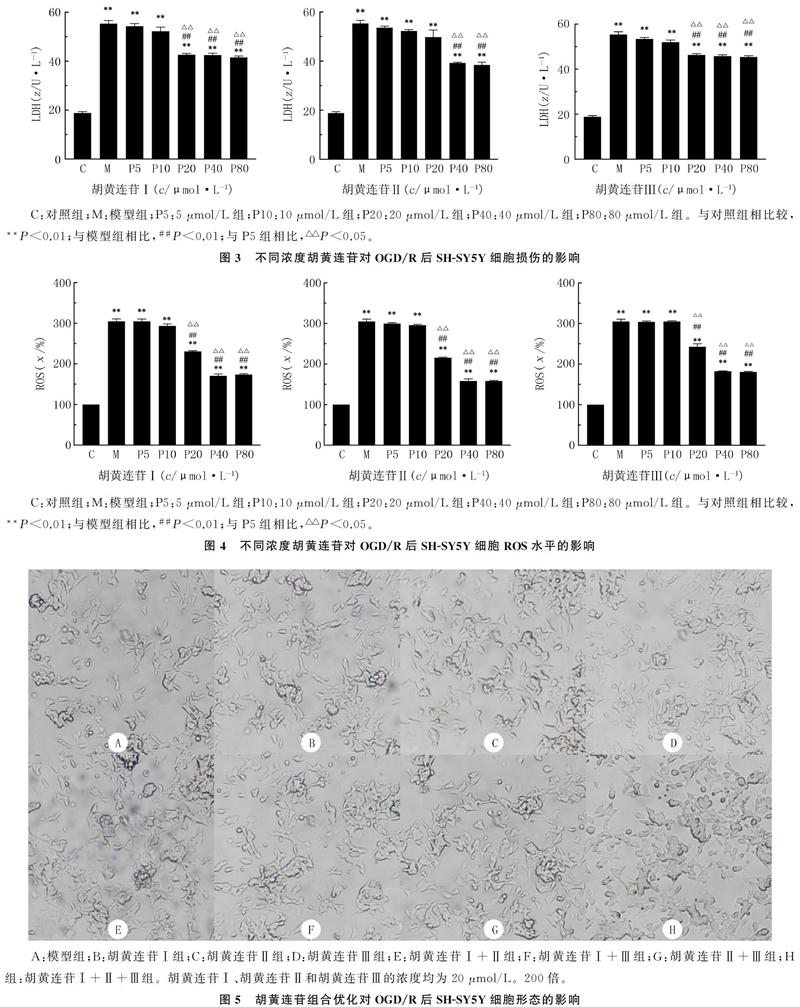
2.2 胡黄连苷对OGD/R后SH-SY5Y细胞存活率的影响
经过OGD/R处理后模型组的细胞存活率较对照组显著下降,差异均有显著性(F=101.9~364.1,t=20.66~38.15,P<0.01);与模型组、胡黄连苷5 μmol/L组相比较,胡黄连苷20、40、80 μmol/L组的细胞存活率明显升高,差异有统计学意义(t=2.71~8.84,P<0.05);但是20、40、80 μmol/L的胡黄连苷对细胞存活率的影响差异无显著性(P>0.05)。根据剂量最小化和疗效最大化的原则,胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ均选择以20 μmol/L为最佳剂量。见图2。
2.3 胡黄连苷对OGD/R后SH-SY5Y细胞损伤程度的影响
与模型组相比,胡黄连苷Ⅰ 20、40、80 μmol/L组和胡黄连苷Ⅲ 20、40、80 μmol/L组SH-SY5Y细胞的LDH释放量明显减少,差异具有统计学意义(F=156.7~252.0,t=8.19~13.78,P<0.01);而胡黄连苷Ⅰ 20、40、80 μmol/L组间和胡黄连苷Ⅲ 20、40、80 μmol/L组间比较差异无显著性(P>0.05)。胡黄连苷Ⅱ 40、80 μmol/L组SH-SY5Y细胞的LDH释放量与模型组相比差异具有统计学意义(F=94.4,t=16.12、16.89,P<0.01)。根据剂量最小化和疗效最大化的原则,胡黄连苷Ⅰ、胡黄连苷Ⅲ均选择以20 μmol/L为最佳剂量,胡黄连苷Ⅱ则选择以40 μmol/L为最佳剂量。见图3。
2.4 胡黄连苷对OGD/R后SH-SY5Y细胞ROS水平的影响
与对照组相比较,OGD/R各组SH-SY5Y细胞的ROS水平均有显著升高(F=385.7~630.0,t=35.41~43.78,P<0.01)。与模型组相比较,胡黄连苷20、40、80 μmol/L组SH-SY5Y细胞的ROS水平均明显降低,差异具有统计学意义(t=12.80~31.39,P<0.01),而胡黄连苷5、10 μmol/L组与模型组比较差异无统计学意义(P>0.05),胡黄连苷40、80 μmol/L组间比较差异无显著性(P>0.05)。根据剂量最小化和疗效最大化的原则,胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ均选择以40 μmol/L为最佳剂量。见图4。
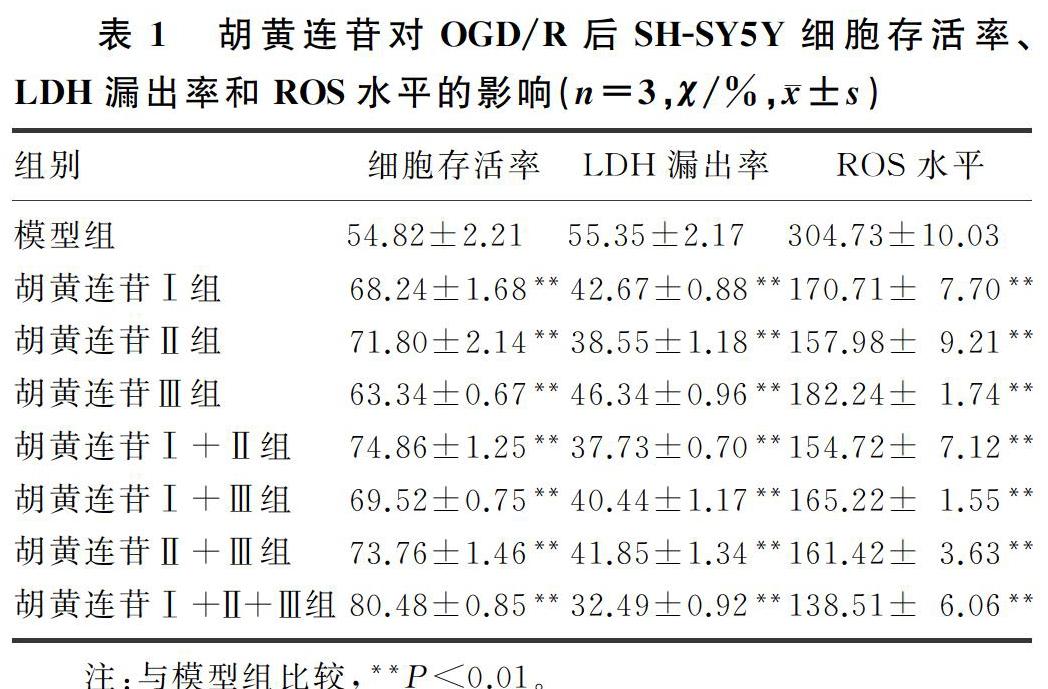
2.5 胡黃连苷组合优化对OGD/R后SH-SY5Y细胞的影响
按照上述细胞形态学、细胞存活率、LDH释放量和ROS水平结果,胡黄连苷Ⅰ最佳剂量分别为20、20、20、40 μmol/L,胡黄连苷Ⅱ最佳剂量分别为20、20、40、40 μmol/L,胡黄连苷Ⅲ最佳剂量分别为20、20、20、40 μmol/L。对各指标进行综合分析,从用药剂量最小化和疗效最大化的角度考虑,选用胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ的最佳剂量为20 μmol/L进行后续实验。
光学显微镜下观察,胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ联合用药组细胞状态最好,较其他组改善更加明显,OGD/R导致的SH-SY5Y细胞损伤明显减轻,虽然细胞突起有所回缩,但是大部分细胞呈长梭形或三角形,贴壁性较其他组好,少部分细胞皱缩变形或脱壁,细胞碎片与模型组相比明显减少。见图5。胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ联合应用组细胞存活率、LDH漏出率、ROS水平结果也优于其他组,差异有统计学意义(F=82.9~186.3,t=2.64~40.17,P<0.05)。见表1。
3 讨 论
脑卒中具有高发病率、高死亡率和高致残率的特点,是导致全球成年人死亡的主要原因之一[22-23]。脑缺血的主要发病机制为脑局部组织血流量减少导致氧气和葡萄糖输送不足以支持细胞代谢需求[24]。脑缺血再灌注损伤是指脑组织缺血后重新获得血流灌注或者氧气供应后产生的一系列损伤和功能障碍,从而引起神经元不可逆性损伤或死亡[25]。研发能够减轻再灌注损伤的神经保护药物至关重要。
胡黄连是我国传统中药,别名割孤露泽、胡连,始载于《唐本草》,功效退虚热、消疳热、清热燥湿、泻火解毒[26]。胡黄连苷的主要有效成分是环烯醚萜苷[27-28]。既往有研究结果表明,在脑缺血再灌注损伤中,胡黄连苷Ⅱ具有抗炎、抗氧化、减轻兴奋性氨基酸的神经毒性、抑制神经元细胞凋亡[29]和神经营养的作用[30]。
研究表明,缺血再灌注损伤与氧化应激反应密不可分[31]。神经元因抗氧化防御能力弱,维持能量稳态能力较低,极易遭受氧化应激损伤[32]。测定神经细胞的存活率是检测神经保护药物作用的常用实验方法[33]。正常情况下LDH不能通过细胞膜,但当细胞膜受到损伤时,漏出到培养液中的LDH增加,故LDH释放量可作为细胞膜完整性的标志物,与细胞损伤程度呈正相关[34]。当体内脑缺血再灌注或体外OGD/R损伤时,细胞的凋亡通常伴随着细胞膜结构的破坏,细胞通透性上升。
本研究结果显示,胡黄连苷对SH-SY5Y细胞OGD/R损伤具有保护作用,且胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ在20 μmol/L时效果最好。胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ各20 μmol/L联合用药,系最佳级联优化组合。胡黄连苷可能通过抗氧化应激发挥神经保护作用,因此胡黄连苷可能是治疗脑缺血再灌注损伤有效且有潜力的药物。胡黄连苷Ⅰ、胡黄连苷Ⅱ、胡黄连苷Ⅲ的最佳剂量及联合应用优化尚未有研究报道,本文结果对减轻缺血低氧引起的神经细胞损伤和保护治疗机制研究具有重要的价值,对进一步了解胡黄连苷的神经保护作用具有重要意义。
[参考文献]
[1]YU Y, WU X Q, PU J N, et al. Lycium barbarum polysaccharide protects against oxygen glucose deprivation/reoxyge-nation-induced apoptosis and autophagic cell death via the PI3K/Akt/mTOR signaling pathway in primary cultured hip-pocampal neurons[J]. Biochemical and Biophysical Research Communications, 2018,495(1):1187-1194.
[2]PANDIAN J D, GALL S L, KATE M P, et al. Prevention of stroke: a global perspective[J]. Lancet, 2018,392(10154):1269-1278.
[3]GBD Lifetime Risk of Stroke Collaborators, FEIGIN V L, NGUYEN G, et al. Global, regional, and country-specific lifetime risks of stroke, 1990 and 2016[J]. The New England Journal of Medicine, 2018,379(25):2429-2437.
[4]CHIU B Y, CHANG C P, LIN J W, et al. Beneficial effect of astragalosides on stroke condition using PC12 cells under oxygen glucose deprivation and reperfusion[J]. Cellular and Molecular Neurobiology, 2014,34(6):825-837.
[5]ZHAO L P, JI C, LU P H, et al. Oxygen glucose deprivation (OGD)/re-oxygenation-induced in vitro neuronal cell death involves mitochondrial cyclophilin-D/P53 signaling axis[J]. Neurochemical Research, 2013,38(4):705-713.
[6]SIES H. Oxidative stress: a concept in redox biology and me-dicine[J]. Redox Biology, 2015,4:180-183.
[7]MANZANERO S, SANTRO T, ARUMUGAM T V. Neuronal oxidative stress in acute ischemic stroke: sources and contribution to cell injury[J]. Neurochemistry International, 2013,62(5):712-718.
[8]BAINES C P. The mitochondrial permeability transition pore and ischemia-reperfusion injury[J]. Basic Research in Cardiology, 2009,104(2):181-188.
[9]WARNE J, PRYCE G, HILL J M, et al. Selective inhibition of the mitochondrial permeability transition pore protects against neurodegeneration in experimental multiple sclerosis[J]. The Journal of Biological Chemistry, 2016,291(9):4356-4373.
[10]VALKO M, LEIBFRITZ D, MONCOL J, et al. Free radicals and antioxidants in normal physiological functions and human disease[J]. The International Journal of Biochemistry & Cell Biology, 2007,39(1):44-84.
[11]ORNELLAS F M, ORNELLAS D S, MARTINI S V, et al. Bone marrow-derived mononuclear cell therapy accelerates renal ischemia-reperfusion injury recovery by modulating inflammatory, antioxidant and apoptotic related molecules[J]. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmaco-logy, 2017,41(5):1736-1752.
[12]LEE J C, WON M H. Neuroprotection of antioxidant enzymes against transient global cerebral ischemia in gerbils[J]. Anatomy & Cell Biology, 2014,47(3):149-156.
[13]JOY K L, RAJESHKUMAR N V, KUTTAN G, et al. Effect of Picrorrhiza kurroa extract on transplanted tumours and chemical carcinogenesis in mice[J]. Journal of Ethnopharma-cology, 2000,71(1-2):261-266.
[14]HAN H, LI Z Q, GAO Z L, et al. Synthesis and biological evaluation of picroside derivatives as hepatoprotective agents[J]. Natural Product Research, 2019,33(19):2845-2850.
[15]GUO Y L, XU X Y, LI Q, et al. Anti-inflammation effects of picroside 2 in cerebral ischemic injury rats[J]. Behavioral and Brain Functions: BBF, 2010,6(7):43-49.
[16]LI S, WANG T T, ZHAI L, et al. Picroside Ⅱ exerts a neuroprotective effect by inhibiting mPTP permeability and EndoG release after cerebral ischemia/reperfusion injury in rats[J]. Journal of Molecular Neuroscience: MN, 2018,64(1):144-155.
[17]LI Q, LI Z, XU X Y, et al. Neuroprotective properties of picroside Ⅱ in a rat model of focal cerebral ischemia[J]. International Journal of Molecular Sciences, 2010,11(11):4580-4590.
[18]PIAO X H, LIU B H, GUO L Y, et al. Picroside Ⅱ shows protective functions for severe acute pancreatitis in rats by preventing NF-κB-dependent autophagy[J]. Oxidative Medicine and Cellular Longevity, 2017, 2017:7085709.
[19]ZHANG S, LU Y M, ZHENG J H, et al. Picroside Ⅰ inhibits asthma phenotypes by regulating Tbet/GATA-3 ratio and Th1/Th2 balance in a murine model of asthma[J]. Tropical Journal of Pharmaceutical Research, 2018,17(9):1777-1782.
[20]PAN J, KONSTAS A A, BATEMAN B, et al. Reperfusion injury following cerebral ischemia: pathophysiology, MR imaging, and potential therapies[J]. Neuroradiology, 2007,49(2):93-102.
[21]TABAKMAN R, JIANG H, LEVINE R A, et al. Apoptotic characteristics of cell death and the neuroprotective effect of homocarnosine on pheochromocytoma PC12 cells exposed to ischemia[J]. Journal of Neuroscience Research, 2004,75(4):499-507.
[22]ZHANG X N, YAN H J, YUAN Y, et al. Cerebral ischemia-reperfusion-induced autophagy protects against neuronal injury by mitochondrial clearance[J]. Autophagy, 2013,9(9):1321-1333.
[23]ZHANG X J, TANG X L, LIU K, et al. Long noncoding RNA Malat1 regulates cerebrovascular pathologies in ischemic stroke[J]. The Journal of Neuroscience, 2017,37(7):1797-1806.
[24]MCCABE C, ARROJA M M, REID E, et al. Animal models of ischaemic stroke and characterisation of the ischaemic penumbra[J]. Neuropharmacology, 2018,134(Pt B):169-177.
[25]HATTORI K, LEE H, HURN P D, et al. Cognitive deficits after focal cerebral ischemia in mice[J]. Stroke, 2000,31(8):1939-1944.
[26]LI P, MATSUNAGA K, YAMAKUNI T, et al. Picrosides Ⅰ and Ⅱ, selective enhancers of the mitogen-activated protein kinase-dependent signaling pathway in the action of neuritogenic substances on PC12D cells[J]. Life Sciences, 2002,71(15):1821-1835.
[27]AN N, WANG D C, ZHU T F, et al. Effects of scrocaffeside A from Picrorhiza Scrophulariiflora on immunocyte function in vitro[J]. Immunopharmacology and Immunotoxicology, 2009,31(3):451-458.
[28]朱仝飞,陈日荣,孙庆文,等. 西藏胡黄连化学成分研究[J]. 中草药, 2017,48(2):263-265.
[29]李琴,郭云良,李震,等. 胡黄连苷Ⅱ对大鼠脑缺血/再灌注损伤Caspase-3和PARP表达的影响[J]. 中国药理学通报, 2010,26(3):342-345.
[30]赵丽,郭云良,李晓丹,等. 胡黄连苷Ⅱ对脑缺血损伤后神经元特异性烯醇化酶表达的影响[J]. 中国药理学通报, 2014,30(2):192-199.
[31]BLOKHINA O, VIROLAINEN E, FAGERSTEDT K V. Antioxidants, oxidative damage and oxygen deprivation stress: a review[J]. Annals of Botany, 2003,91(2):179-194.
[32]BOLAOS J P, ALMEIDA A. The pentose-phosphate pathway in neuronal survival against nitrosative stress[J]. IUBMB Life, 2010,62(1):14-18.
[33]CHE Q, WANG W, DUAN P, et al. Downregulation of miR-322 promotes apoptosis of GC-2 cell by targeting Ddx3x[J]. Reproductive Biology and Endocrinology: RB&E, 2019,17(1):63.
[34]HILLION J A, LI Y X, MARIE D, et al. Involvement of Akt in preconditioning-induced tolerance to ischemia in PC12 cells[J]. Journal of Cerebral Blood Flow & Metabolism, 2006,26(10):1323-1331.
(本文編辑 马伟平)
