
意大利苍耳与乌拉尔甘草种间竞争能力比较
2021-10-09 01:19徐怀秀
生态学报 2021年16期
徐怀秀,马 淼
新疆植物药资源利用教育部重点实验室,石河子大学生命科学学院, 石河子 832003
生物入侵是当前世界性的生态环境问题[1],随着全球化进程加快,入侵物种严重威胁到了各国生态系统的完整性[2]。其中农田生态系统中植物群落结构简单,稳定性较低,而入侵植物大多具有强大的竞争力,这使得农田生态系统往往最易受到外来植物的成功入侵[3]。一些与作物具有相似生态位的入侵植物,会通过种间竞争来获得生存优势,并对作物的生长、繁殖和产量积累产生强烈的负面影响[4- 5]。
意大利苍耳(XanthiumitalicumMoretti)为菊科(Compositae)苍耳属(Xanthium)一年生草本植物,原产于北美洲,现广泛分布于东、西半球中纬度地区,已成为世界性入侵杂草[6]。1991年在我国北京首次发现,目前已经广泛分布至辽宁、北京、山东、河北、陕西、新疆等地,并且有扩大蔓延至全国各地的趋势[7- 8]。其中意大利苍耳在新疆的危害最为严重,目前在新疆已扩散至伊犁、博州、阿勒泰、塔城、石河子、昌吉、乌鲁木齐等地区,已占据了农田、草原、林缘、沟渠及水库浅水区等多种生境[9]。对新疆的农业生产和畜牧业发展都产生了极大危害[10- 11]。
乌拉尔甘草(GlycyrrhizauralensisFisch)为豆科(Leguminosae)甘草属(Glycyrrhiza)多年生草本植物,其根及根状茎是西北地区的特色道地性药材[12],其中含有的甘草酸和甘草苷等黄酮类活性成分,具有抗病毒、抗癌、抗过敏、抗炎等功效[13]。同时,乌拉尔甘草的茎叶部分也是优良的饲草,被广泛应用于畜牧业[14]。极高的药用价值及经济价值使其成为中国西北地区大面积种植的一种经济作物。本课题组前期在新疆的野外调查中发现,意大利苍耳已经开始向乌拉尔甘草农田扩张,与乌拉尔甘草形成混生的格局。研究表明,外来入侵植物进入新生境后会与本地植物通过抢占空余生态位或通过种间竞争获得优势[15- 16],其中种间竞争是入侵植物排挤本地种的一种重要策略,其主要是基于资源的竞争性利用[5]。影响植物种间竞争结果的因素包括种间竞争能力的遗传差异,资源的有效性,生态位相似性,共生微生物,空间分布格局,竞争期长短等[17]。入侵植物意大利苍耳具有较高的生长速率,易于形成高密度单优势种群落,而农田生境通常具有较好的水和营养资源,这使得农田中意大利苍耳的株高和冠幅能够分别达到2 m和1.6 m[9, 18],高大的个体和庞大的冠幅使意大利苍耳对农田作物产生严重的遮蔽,从而造成农田作物的严重减产[4]。在乌拉尔甘草农田中,甘草的株高通常不到1 m,在与意大利苍耳混生的群落中位于意大利苍耳的下层,这可能使得意大利苍耳在对光资源的竞争中占有优势地位。此外,意大利苍耳旺盛的生长,势必会加大对土壤中水分和矿质营养的获取,而这也可能会加剧其与农田中的甘草之间对于资源的竞争。以往的研究更多关注的是意大利苍耳对入侵地自然生态系统所造成的影响[6,8,10, 19],而其对农田的入侵风险及后果的评估研究较少,其与甘草属植物之间的竞争关系也不明确,关于入侵植物意大利苍耳与乌拉尔甘草的种间竞争会对乌拉尔甘草的生长及品质会产生怎样的影响尚不清楚。因此,本研究采用取代试验法,在受控条件下,模拟栽培乌拉尔甘草常规水肥管理条件,开展了意大利苍耳与乌拉尔甘草种间竞争关系及竞争结果的研究,以期为意大利苍耳对乌拉尔甘草农田的入侵能力及入侵后果的评估提供试验依据。
1 材料与方法
1.1 试验区概况
试验地点位于新疆石河子市石河子大学(44°31′10″N, 86°06′94″E),海拔约450 m,属典型的大陆性干旱气候,光照充足,降水少蒸发大。年均降水量180 mm左右,年均蒸发量1820 mm左右,年均气温为7.2℃,年日照时数2721—2818 h,无霜期约147 d。
1.2 试验材料
试验所用的意大利苍耳成熟种子于2018年10月采自新疆石河子市东郊(44°30′01″N, 86°12′35″E);乌拉尔甘草的成熟种子于2018年9月采自新疆尉犁县(41°38′83″N, 86°28′73″E),种子自然阴干后置于4℃保存。
1.3 试验方法
1.3.1种子预处理
为保证种子萌发的同步性,选取大小一致、成熟饱满的意大利苍耳种子,播种前在室温下置于蒸馏水中浸泡12 h;选取大小一致、成熟饱满的乌拉尔甘草种子,用98%的硫酸溶液浸泡约0.5 h,用流水冲洗至种子表面无硫酸残留,在室温下用蒸馏水浸泡12 h。
1.3.2试验设计
于2019年4月15日将经过水浸泡处理的意大利苍耳和乌拉尔甘草种子播种于育苗盆中,以沙土作为栽培基质,播种深度为1 cm,置于户外开阔地,每天用称重法补水以保持土壤相对湿度为70%左右,待幼苗长出4枚真叶时选长势一致的幼苗并将其移栽于直径为22 cm,高度为20 cm的塑料花盆中。根据甘草农田高产管理栽植技术[20],其栽植密度通常为120万株/ hm2,与盆栽面积进行换算,确定本研究中总种植密度为4株/盆。采用取代试验法[17, 21],设置单种种植和混种种植两种模式,共计三种处理,即单种种植意大利苍耳,单种种植乌拉尔甘草以及意大利苍耳与乌拉尔甘草混合种植,单种种植指每盆单独种植4株意大利苍耳(即意大利苍耳∶乌拉尔甘草=4∶0)或每盆单独种植4株乌拉尔甘草(即意大利苍耳:乌拉尔甘草=0:4),混种种植时每盆种植意大利苍耳和乌拉尔甘草各2株(即意大利苍耳:乌拉尔甘草=2:2),四株个体在花盆中呈“十”字形均匀分布,两种植物相同排列,具体种植方式见图1。

图1 意大利苍耳和乌拉尔甘草栽植示意图Fig.1 The planting diagram of Xanthium italicum and Glycyrrhiza uralensis
栽培基质为沙土(全氮0.268 g/kg、全磷0.0855 g/kg、全钾5.72 g/kg、速效氮43.59 mg/kg、速效磷4.1 mg/kg、速效钾119.09 mg/kg、有机质5.81 g/kg)。使用称重法,每天早晚补水,保持70%的土壤相对湿度;模拟甘草农田土壤肥力水平[22]分2次(2019年5月21日和2019年5月28日)对各个花盆进行施肥(1.4492 g尿素、1.9354 g过磷酸钙、1.7108 g硫酸钾)。每种处理重复6次,计18盆。所有花盆都随机摆放在开阔地,每隔10 d随机调换一次各花盆的位置,以消除光照和边际效应的影响。于2019年9月1日意大利苍耳生育期结束(即果实成熟)后,收获全部植株。
1.4 研究指标及方法
1.4.1形态学参数的测定
植株收获后,用水将植株的根部冲洗至无浮土,测量所有植株的株高、冠幅及乌拉尔甘草的根状茎长度;统计意大利苍耳的种子数量和乌拉尔甘草的根状茎条数与根瘤菌数量;用根系扫描仪(WinRHIZO La 2400)测量乌拉尔甘草的根总长度、根总表面积及根平均直径。
1.4.2生物量参数的测定
将每株植株的根、茎、叶、果实(意大利苍耳)、根状茎(乌拉尔甘草)进行分离,置于60℃烘箱中烘干至恒重,称量各部分生物量并计算总生物量。
1.4.3甘草酸含量的测定
将烘干的乌拉尔甘草的根及根状茎混合粉碎成粉末,过80目筛。分别精确称取单种处理组和混种处理组的粉末各0.50 g,置于10 mL离心管中,加入提取液5 mL(甲醇: 水: 36%冰醋酸= 71: 28: 1),用超声波提取仪超声提取30 min,用高速离心机(8000 r/min)离心10 min,吸取上清液,沉淀物再加入5 mL提取液,超声提取30 min,用高速离心机(8000 r/min)离心10 min,吸取上清液,并与第一次的上清液混匀,上清液经0.45 μm微孔滤膜过滤于安捷伦进样瓶中。使用高效液相色谱仪(Agilent 1200; Agilent Technologies, CA, USA)测量甘草地下器官(根及根状茎)的甘草酸含量[23]。
1.4.4竞争关系
分别计算这两种植物的相对产量(Relative Yield,RY)[21]、相对竞争强度(Relative Competition Intensity, RCI)[24]以及竞争攻击系数(Aggressivity,A)[25],以确定两物种间的竞争关系,其公式为:
RYa=Yab/(Zab×Yaa) 或RYb=Yba/(Zba×Ybb)
RCIa=(Yaa-Yab)/Yaa或 RCIb=(Ybb-Yba)/Ybb
Aa=Yab/(Yaa×Zab)-Yba/(Ybb×Zba) 或Ab=Yba/(Ybb×Zba)-Yab/(Yaa×Zab)
式中,a、b分别代表意大利苍耳和乌拉尔甘草,Yab代表混合种植处理组中意大利苍耳的单株生物量,Yaa代表单独种植时意大利苍耳的生物量,Yba代表混合种植处理组中乌拉尔甘草的单株生物量,Ybb代表单独种植时乌拉尔甘草的生物量,Zab为混合种植体系中意大利苍耳所占的比例,Zba为混合种植体系中乌拉尔甘草所占的比例。
当RY<1时,表示种间竞争大于种内竞争;RY> 1时,表示种内竞争大于种间竞争;RY=1时,表示种间竞争等于种内竞争。
当RCI值位于0—1之间时,RCI值越大,说明该物种在混种环境中产量越低,另一植物对它的影响越大,且该物种在混种群落中的竞争能力越差;当RCI=1时说明该物种已被排斥出去;当RCI=0时,说明该物种对混种的其他物种在产量上没有影响。当RCI<0时,表明一种植物的存在能够促进该植物的生长,促进其产量。
当Aa> 0时,表明物种a的竞争能力大于物种b;当Aa= 0时,表明物种a与物种b的竞争力相同,当Aa< 0时,表明物种a的竞争能力小于物种b。
1.5 数据处理
使用SPSS 19.0软件进行数据的统计分析,用单因素方差分析(one-way ANOVA)和非参数检验(nonparametric tests)分析每种植物在单种处理和混种处理下的形态学性状、生物量及甘草的地下器官甘草酸含量的差异显著性,使用T检验(T-Test)比较RY与1,以及RCI、A与0之间的差异显著性,使用origin8. 5软件进行作图。
2 结果
2.1 种植模式对意大利苍耳和乌拉尔甘草生长的影响
种植模式对意大利苍耳的株高、冠幅及种子数量均具有极显著影响(P<0.01)。混种处理下意大利苍耳的株高、冠幅及种子数量相比单种处理分别增加了13%、27%和56%(图2)。

图2 种植模式对意大利苍耳生长和生物量积累的影响(均值±标准误) Fig.2 Effects of planting pattern on growth and biomass accumulation of Xanthium italicum (mean±SE)不同大写字母表示单种与混种处理在1%水平上差异显著
种植模式也极显著地影响了乌拉尔甘草的株高、冠幅、根总长度、根总表面积、根平均直径、根状茎条数、根状茎长度和根瘤数(P<0.01)(表1)。混种处理组中乌拉尔甘草的株高和冠幅相比单种处理组分别下降了35%和45%。单种处理下单株乌拉尔甘草的平均根状茎条数为3条,根状茎总长度平均为1.6 cm,而混种处理组中的乌拉尔甘草均未生长出根状茎。混种处理组中乌拉尔甘草的根总长度、根总表面积、根平均直径及根瘤数相比单种处理分别下降了55%、63%、19%和76%。

表1 种植模式对乌拉尔甘草的生长的影响(均值±标准误)
2.2 种植模式对意大利苍耳和乌拉尔甘草生物量分配的影响
种植模式对意大利苍耳的根、茎、叶、果实和总生物量均具有极显著影响(P<0.01)。意大利苍耳的根、茎、叶、果实及总生物量在混种处理下相比单种处理,分别增长了84%、73%、84%、73%和77%(图2)。
种植模式也显著影响了乌拉尔甘草的根状茎生物量(P=0.014),极显著地影响其根、茎、叶及总生物量的积累(P<0.01)。单种条件下乌拉尔甘草的根状茎生物量为0.016 g,而混种处理下乌拉尔甘草未生长出根状茎,其根、茎、叶及总生物量相比单种处理分别下降了72%、80%、65%和71%(图3)。
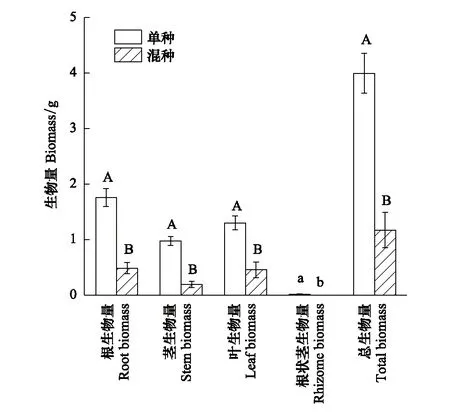
图3 种植模式对乌拉尔甘草生物量的影响(均值±标准误) Fig.3 Effects of planting pattern on the biomass of Glycyrrhiza uralensis (mean±SE)不同大写字母表示单种与混种处理在1%水平上差异显著,不同小写字母表示单种与混种处理在5%水平上差异显著

图4 种植模式对乌拉尔甘草地下器官甘草酸含量的影响(均值±标准误)Fig.4 Effects of planting pattern on glycyrrhizic acid content in underground organ of Glycyrrhiza uralensis (mean±SE)
2.3 种植模式对乌拉尔甘草甘草酸含量的影响
种植模式极显著地影响了乌拉尔甘草地下器官甘草酸含量的积累(P<0.01)。混种处理组中乌拉尔甘草的甘草酸含量比单种处理组下降了63%(图4)。
2.4 意大利苍耳和乌拉尔甘草的竞争关系
意大利苍耳的相对产量(RYa)大于1,表明意大利苍耳的种内竞争大于来自乌拉尔甘草的种间竞争;其相对竞争强度(RCIa)小于0,表明与乌拉尔甘草混种会有利于意大利苍耳生物量的积累;其竞争攻击系数(Aa)大于0,表明混种的意大利苍耳的竞争能力大于乌拉尔甘草(图5)。
乌拉尔甘草的相对产量(RYb)小于1,表明乌拉尔甘草的种内竞争小于来自意大利苍耳的种间竞争;其相对竞争强度(RCIb)在0—1之间,表明意大利苍耳对乌拉尔甘草的影响较大且乌拉尔甘草在混种群落中竞争能力较弱;其竞争攻击系数(Ab)小于0,表明混种的乌拉尔甘草竞争能力小于意大利苍耳(图5)。
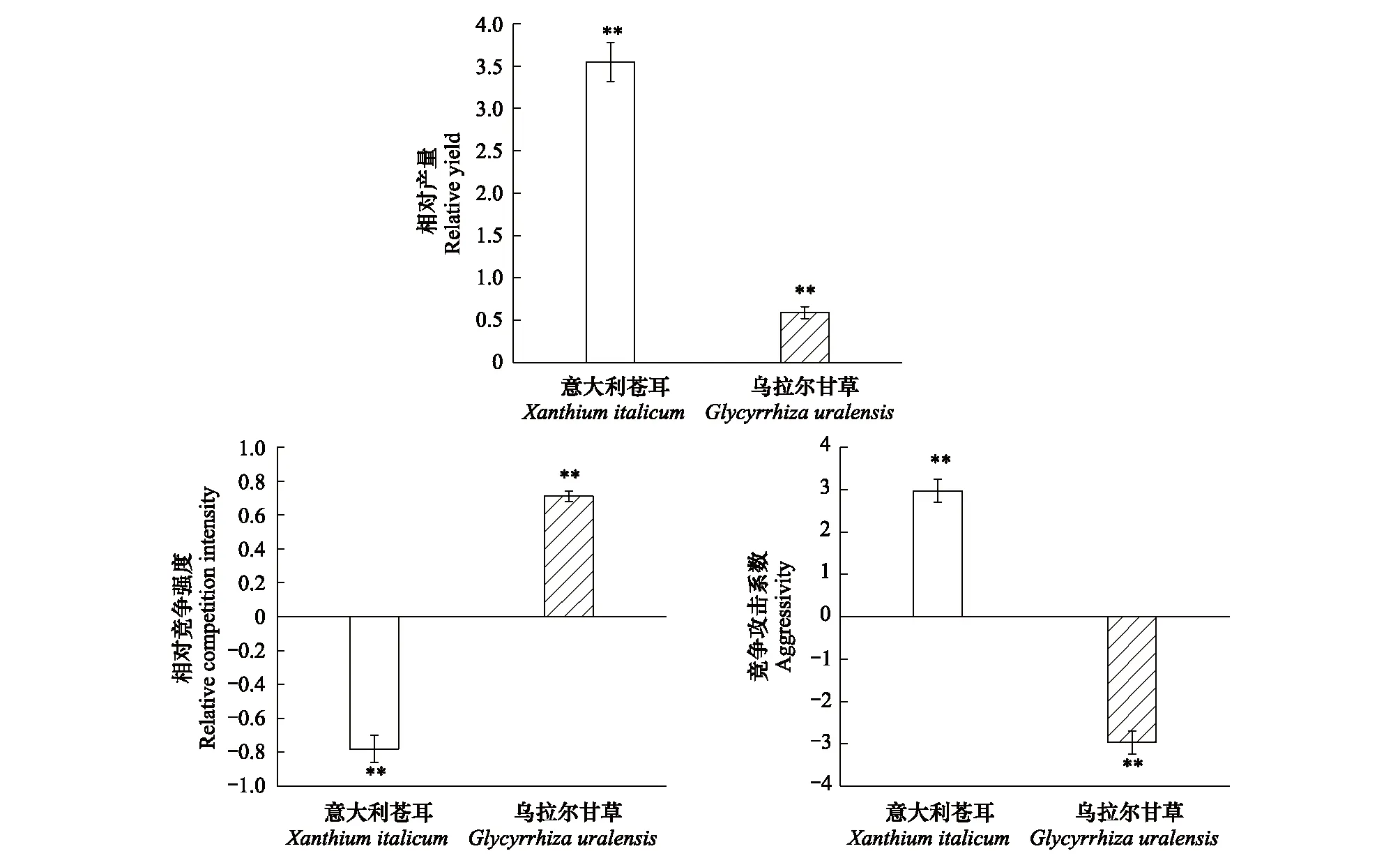
图5 不同种植模式下该两种植物的竞争关系(均值±标准误)Fig.5 Competitive relationship of the two species based on different planting pattern (means±SE) **表示竞争指标与假设检验值之间(即RY与1比较,RCI和A分别与0比较) 存在极显著差异,(单一样本T检验:P<0.01, n=12)
3 讨论
外来植物与本地植物之间的竞争关系是入侵生态学的重要研究内容之一,外来植物能否成功入侵,主要取决于该植物自身的入侵力与生境的可入侵性[26]。而该植物自身入侵力主要取决于物种本身的生物学特性,其中就包括该植物的种间竞争能力[27- 28]。不同的物种具有不同的竞争策略,而植株的大小特征是预测植物相对竞争力的最佳指标,生物量是其中最重要的参数[29]。在本研究中,与单种相比,混种能够显著提高意大利苍耳根、茎、叶及果实的生物量,这意味着混种处理会有利于该入侵植物的生物量积累,从而提高其相对竞争力[30- 31]。我们发现与单种模式相比,在混种处理中意大利苍耳根和叶的生物量增加幅度最大,这意味着存在竞争作用时,意大利苍耳的生物量分配会趋向于增加根和叶器官的生长,发达的根系使其能更高效地吸收土壤中的水分和无机营养,而发达的叶片则意味着更大的光合面积,这有助于其制造出更多的光合产物,使其具有更大的营养器官,从而提高其自身的入侵力[32]。混种处理中意大利苍耳的茎生物量也显著增加,在与乌拉尔甘草混生时,意大利苍耳能够生长出更高的个体和更大的冠幅,使其在地上空间占领方面具有更明显的优势,造成物种间竞争的不平衡[33],进而有利于该入侵植物对甘草农田的成功入侵。研究表明,意大利苍耳的种子具有良好的越冬性能,其在各种极端条件下越冬依旧能够保持较高的萌发率[11]。本研究中,混种使意大利苍耳的种子数量显著增加,这意味着入侵甘草地后,意大利苍耳能够产生更多的种子,良好的越冬性能及数量巨大的种子使其有性繁殖能力得到显著提高,于是在下一个生长季节,被意大利苍耳入侵的乌拉尔甘草农田中将会出现更多的意大利苍耳个体,而这也将加剧乌拉尔甘草遭受到的意大利苍耳的竞争抑制威胁。
生境的可入侵性主要取决于非生物环境和生物环境对入侵植物的抵抗力[34],农田生境作为一种高强度的人为扰动环境,其资源可用量会受人为扰动发生改变[35],而这种高强度的扰动通常都利于外来物种的成功入侵[36]。因此,农田生境的可入侵性,则主要取决于生物环境对入侵植物的抵抗力,其中作物与入侵植物的种间竞争就是一种重要的抵抗策略[37]。在本研究中,混种处理中的乌拉尔甘草受到意大利苍耳强烈的竞争抑制,各部分生物量的积累均显著降低。相对于单种处理,混种处理中乌拉尔甘草的株高、冠幅及根系的生长参数均呈现大幅的下降,这些营养器官的生长受到显著抑制,必然会对乌拉尔甘草叶片的光捕获能力和根系对土壤水分及营养资源的获取能力产生极其不利的影响。这意味着在该两种植物对主要资源的竞争利用中,乌拉尔甘草处于不利的地位,对意大利苍耳的入侵表现出较弱的抵抗力。同时意大利苍耳还强烈地抑制了甘草根状茎的发育,这使得乌拉尔甘草的克隆繁殖潜力被显著降低。而甘草酸是中药材甘草的主要活性成分[38],药用植物有效成分的含量会受伴生植物种类的强烈影响[39]。本研究结果显示,混种处理组中乌拉尔甘草地下器官甘草酸含量极显著地低于单种处理组的乌拉尔甘草,这表明意大利苍耳能够显著降低中药材甘草的品质。综合来看,与意大利苍耳混生情况下,乌拉尔甘草的个体生长、克隆繁殖能力以及药材甘草的产量与品质均受到强烈的抑制。
共生微生物是影响植物竞争的一种重要因素之一[17]。根瘤菌可以与豆科植物的根形成共生体系,这种共生体系是生物固氮中最重要的一种类型[40]。本研究中发现,与单种相比,混种处理组中乌拉尔甘草根系的根瘤数量明显降低,其数量仅是单种处理组的四分之一左右,这表明与意大利苍耳的混种处理会强烈抑制根瘤菌与乌拉尔甘草根共生体系的形成。研究表明,混生情况下根瘤菌的缺失对豆科植物的影响明显大于其他植物[41],但本研究中甘草根瘤数量减少的内在机制尚不明确,需进行更深入的研究。但从长远来看,这种根瘤发育的受阻和根瘤数量的减少,势必会对农田土壤的氮素水平产生负面的影响。
植物的相对产量、相对竞争强度以及竞争攻击系数是解释物种间竞争关系的重要指标[42]。在本研究中,相对产量的结果表明,意大利苍耳受到邻株同种其他个体的种内竞争压力大于来自邻株乌拉尔甘草的种间竞争压力,而乌拉尔甘草受到邻株同种其他个体的种内竞争压力则小于来自邻株意大利苍耳的种间竞争压力。相对竞争强度结果显示,混种模式对意大利苍耳的生长有显著的促进作用,这可能是由于邻株的乌拉尔甘草具有良好的固氮作用[40],因此与意大利苍耳单种模式相比,混种模式可能具有更高的土壤氮素水平,而入侵植物通常能够比本地植物具有更高的氮资源利用能力[43],从而可能影响到两种植物间的竞争关系。竞争攻击力系数的数据表明,意大利苍耳的竞争能力大于乌拉尔甘草,在二者混生的群落中,意大利苍耳将会在竞争中占据优势地位。
综上所述,在与乌拉尔甘草混合种植时,意大利苍耳的营养生长和生殖生长均得到了极显著的促进,使其个体比单种处理组长得更高大,并能产生更多的种子。这不仅显著地抑制了乌拉尔甘草的生长,而且强烈削弱了乌拉尔甘草凭借根状茎的克隆繁殖能力,并使乌拉尔甘草的药材产量显著降低,同时还强烈抑制了甘草酸的形成和积累,从而消弱了甘草药材的品质。该药材产量和品质的大幅下降必然会给农户造成巨大的经济损失。甘草茎和叶生物量的显著降低也势必造成家畜饲草产量的下降,从而危及当地畜牧业的健康发展。
猜你喜欢
草业学报(2022年8期)2022-11-23
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国现代医生(2019年5期)2019-04-25
蔬菜(2018年5期)2018-05-17
现代兵器(2016年4期)2016-04-27
杂草学报(2015年2期)2016-01-04
绿色科技(2015年6期)2015-08-05
江苏农业科学(2014年7期)2014-09-02
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
