
鸟类取食和果实成熟度对日本珊瑚树种子萌发的影响
2021-10-09 01:42:00沈世峰任星榕杨心语邓仕明
湖北民族大学学报(自然科学版) 2021年3期
沈世峰,任星榕,韩 慧,杨心语,邓仕明
(1.生物资源保护与利用湖北省重点实验室(湖北民族大学),湖北 恩施 445000;2.恩施州特色植物资源种质工程技术研究中心(湖北民族大学),湖北 恩施 445000;3.湖北民族大学 林学园艺学院,湖北 恩施 445000)
种子传播对于植物群落构建、种群更新与扩散[1]及植物入侵[2]均具有重要作用.食果动物常通过取食与排泄、无意识携带等方式帮助种子传播[3].研究表明,食果鸟对肉果的取食,不仅能促进种子传播[4],从而降低子代与母株及子代之间的竞争[5],还能提高成苗率[6].
食果鸟对肉果的取食方式可分为啄食和吞食两种[7].经食果鸟啄食过的果实,常被去除部分或全部果肉,种子多不经肠道而直接掉落于树下;被吞食的果实,其肉质果肉会被鸟的消化道完全消化掉,相对难消化的种子(核果类常带有坚硬的内果皮)会随鸟的粪便排出.已有的研究表明,经过鸟类肠道随粪便排泄而出的种子往往更易萌发[8].这可能是因为消化道去除了含有萌发抑制物的果肉[9],或者消化道对果/种壳有划破作用[10],或者两种作用兼而有之.
在肉果发育后期,外果皮会因叶绿素逐渐分解和花青素的合成而发生颜色变化,通常由绿色变成红、黄、橙、黑等颜色[11],一方面昭示着果实的成熟度,另一方面也昭示着种子的成熟状态[12].果实颜色影响食果鸟对果实的取食[13-15],其中红色和黑色果实往往是食果鸟的首选[16].果实颜色(或成熟度)与食果鸟的取食偏好对种子传播和萌发具有怎样的影响?目前鲜见研究.这方面的研究对于植物物种保护和生态管理具有重要意义.
日本珊瑚树(Viburnumodoratissimumvar.awabuki(K.Koch) Zabel)为五福花科(Adoxaceae)荚蒾属常绿灌木或小乔木,是珊瑚树(V.odoratissimum)的一个变种[17-19],具有较高园林观赏价值.其果实为卵圆形肉质核果,呈圆锥果序,成熟时先由绿色变为红色,而后逐渐转变成紫黑色[20],果实量大且果期较长,成熟期间常吸引多种食果鸟取食.目前关于日本珊瑚树种子休眠与萌发的相关研究较少,仅有少数关于其原变种的(珊瑚树).原变种珊瑚树的种子具有非深度简单上胚轴形态生理休眠[21-22],而对于日本珊瑚树种子的研究仅知其具有低温休眠特性,赤霉素和冷层积处理虽能打破其休眠,但萌发率都不是很高[23].
1 材料与方法
1.1 试验材料
日本珊瑚树果实于2018年9月15日采自湖北民族大学校园内(N30°17′57.66″,E109°30′28.98″,海拔450 m)种植的至少30株壮年植株.采回的果实按果皮颜色分为红色和紫黑色两组.
1.2 食果鸟取食观察
参照林佩蓉等[13]的方法对食果鸟及其取食日本珊瑚树果实情况进行观察.观察于9月开始,时值刚刚进入果熟期,果实开始由绿色向红色转变,直至果实几乎全部脱落.观察时间为每天上午7:30-8:30和下午17:20-18:30,以肉眼观察为主,辅以长焦望远镜观察和长焦照相机拍照.所拍照片用于鉴定食果鸟种类.观察食果鸟及其取食行为的同时,对掉落树下的果实(或种子)和鸟粪中的种子进行观察和收集,以明确食果鸟取食对果实和种子物理结构的影响.
1.3 去除果肉处理和划破内果皮处理
参照Vélez等[24]的方法,将采回的红色和紫黑色果实分别取一部分人工去除果肉,以获得仅具有坚硬内果皮的种子,再从中分别随机取一部分已去除果肉的种子,用解剖刀在萌发孔一端划破内果皮,同时避免弄伤里面真正种子,随后进行萌发测试,以检测去除果肉和划破内果皮对种子萌发的影响.
1.4 萌发测试
参照Vélez等[24]的方法,将完整果实(对照)、人工去除果肉种子及去除果肉后划破内果皮的种子均匀播于盛有湿润珍珠岩的培养皿(d=9 mm)内,然后置于15 ℃光照培养箱(每天12 h光照/12 h黑暗交替处理,光照强度为121 μmol/(m2·s)内进行培养.每个处理设置3个重复,每个重复25粒种子,每隔4 d观察并记录一次萌发情况.整个萌发测试持续124 d,期间适时补水,以胚根突破种皮大于等于2 mm作为萌发标准,按下面公式分别计算萌发率、萌发势和萌发指数.
式中:G为萌发率,GP为萌发势,GI为萌发指数,N1为试验结束时正常萌发的种子数,N2为供试种子数,N3为日平均发芽数达到最高时的正常萌发种子数,Gt为第t天的种子萌发数,Dt为种子萌发天数.
1.5 数据处理与分析
统计分析和做图由R 3.6.0软件完成.相同处理条件下两种成熟度种子开始萌发时间之间的方差分析采用独立样本t检验;同一成熟度种子不同处理间的开始萌发时间之间的方差分析采用单因素方差分析,并使用最小显著性差异法(least significant difference,LSD)进行验后多重比较;两种成熟度果实经三种处理后种子的萌发率、萌发势和萌发指数之间的差异采用多因素方差分析,并使用LSD方法进行验后多重比较,若交互作用显著则进一步进行简单效应分析.为了保证方差齐性,将萌发率数据进行平方根反正弦转化[25]后再进行方差分析.文中所有数据均以平均值±标准误表示,小数点后面保留两位有效数字.
2 结果与分析
2.1 食果鸟取食观察
日本珊瑚树果实成熟时颜色先由绿色变成红色,再由红色转变成紫黑色.两种颜色的果实均已达生理成熟.观察发现,日本珊瑚树下掉落的种子,有的带果肉,有的不带果肉,其中带果肉的果实多为紫黑色.同时,共观察到4种食果鸟取食日本珊瑚树果实,如表1所示.
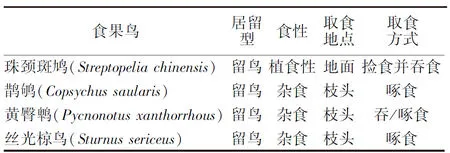
表1 食果鸟取食日本珊瑚树果实相关信息Tab.1 Information on frugivorous bird feeding on V.odoratissimum var.awabuki fruits
2.2 果实成熟度和模拟食果鸟取食对种子萌发的影响
对于红色果实而言,去除果肉处理和去除果肉后划破内果皮处理的种子的萌发率、萌发势和萌发指数均与完整果实(对照组)无显著差异(P>0.05,如图1所示).对于紫黑色果实而言,去除果肉处理显著提高了种子的萌发率、萌发势和萌发指数(P<0.05),而去除果肉后划破内果皮处理的种子的萌发率和萌发指数与完整果实和去除果肉处理均无显著差异(P>0.05),并且去除果肉后划破内果皮处理的种子的萌发势与完整果实也无显著差异(P>0.05),并显著低于去除果肉处理(P<0.05,如图1所示).

红色果实的种子萌发数据间的方差分析结果用小写英文字母标记;紫黑色果实的种子萌发数据间的方差分析结果用大写英文字母标记;相同处理的红色果实和紫黑色果实的种子萌发数据间方差分析结果用星号标记;标有相同小写或大写英文字母的数据间在P=0.05水平上差异不显著;单个星号“*”表示在P=0.05水平上具有显著性差异;两个星号“**”表示在P=0.01水平上具有极显著性差异;“NS”表示在P=0.05水平上无显著性差异.图1 去除果肉和去除果肉后划破内果皮处理对红色和紫黑色两种成熟度果实中种子萌发率(A)、萌发势(B)和萌发指数(C)的影响Fig.1 Effects of pulp-removed and endocarp-scarified treatments on germination (A),germination potential (B) and germination index (C) of seeds in red and purple-black fruits
对于完整果实和去除果肉后划破内果皮处理而言,红色果实的种子的萌发率和萌发势分别显著(P<0.05)和极显著(P<0.01)高于紫黑色果实,但两种成熟度果实的萌发指数间并无显著差异(P>0.05);而对于去除果肉处理而言,两种成熟度果实的种子萌发率、萌发势和萌发指数间均无显著差异(P>0.05,如图1所示).
红色和紫黑色果实经不同处理后种子开始萌发所需培养时间如表2所示,由表2可知,去除果肉后划破内果皮处理的红色果实的种子开始萌发时间最早,平均需培养(53.33±1.33)d,而完整紫黑色果实的种子开始萌发时间最晚,平均需培养(80.00±4.00)d.红色和紫黑色果实经相同的处理后,种子开始萌发所需培养时间无显著差异(P>0.05).对于红色果实而言,与完整果实相比,去除果肉后划破内果皮处理显著缩短了种子开始萌发前的培养时间(P<0.05),而去除果肉处理则与完整果实无显著差异(P>0.05).对于紫黑色果实而言,与完整果实相比,去除果肉处理和去除果肉后划破内果皮处理均显著缩短了种子开始萌发前的培养时间(P<0.05),而两种处理间无显著差异(P>0.05).
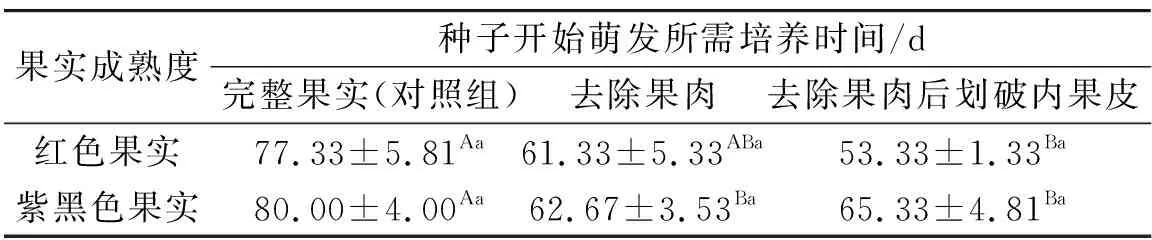
表2 红色和紫黑色果实经不同处理后种子开始萌发所需培养时间Tab.2 Culture time for seeds in red and purple-black fruits starting to germinate
2.3 果实成熟度和模拟食果鸟取食处理之间的交互作用分析
果实成熟度与处理方式之间的交互作用分析如表3所示,成熟度对种子萌发率和萌发势具有极显著影响(P<0.01),对萌发指数具有显著影响(P<0.05);处理方式对萌发率、萌发势和萌发指数均无显著影响 (P>0.05),成熟度和处理方式间也无显著的交互作用(P>0.05);仅在萌发势方面,成熟度和处理方式间存在显著的交互作用 (P<0.05).
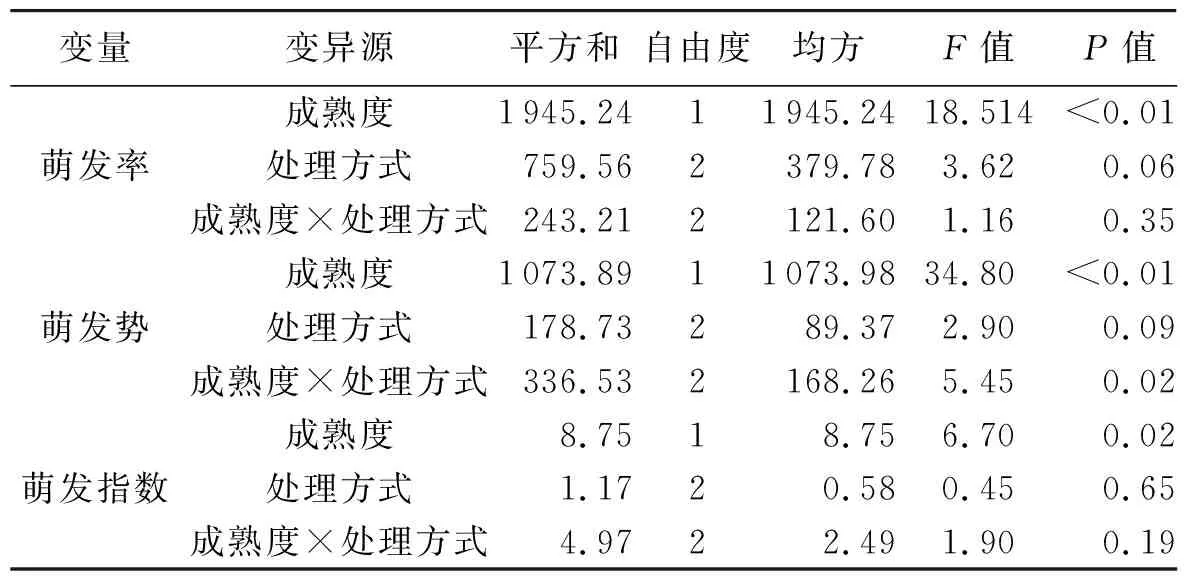
表3 果实成熟度与处理方式之间的交互作用分析Tab.3 Analysis of the interaction between the fruit maturity degree and treatment method
对交互作用所做的简单效应分析表明,对于种子的萌发势,在成熟度与完整果实间,成熟度与去除果肉后划破内果皮处理间,以及处理方式与紫黑色果实间均存在极显著的交互作用效应(P<0.01),而在成熟度与去除果肉处理间及处理方式与红色果实间不存在显著的交互作用效应(P>0.05,如表4所示).
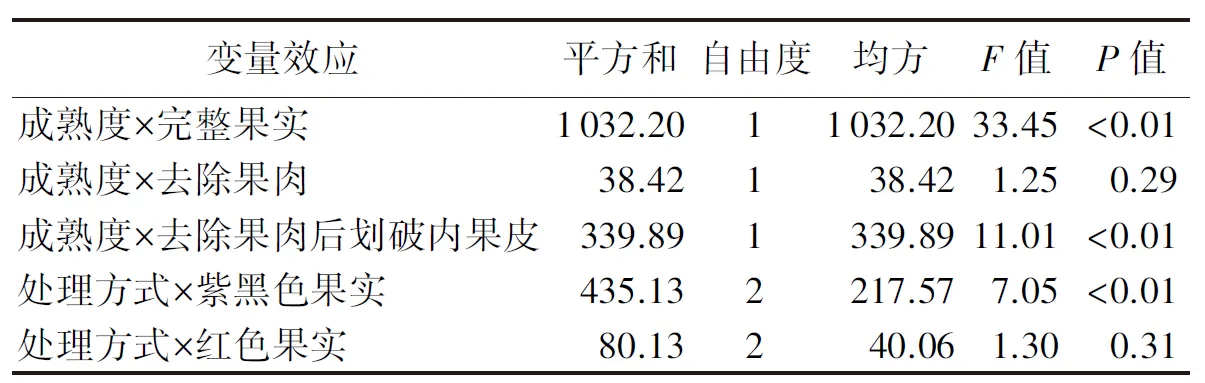
表4 果实成熟度与处理方式在影响种子萌发势方面的交互作用的简单效应分析Tab.4 Simple effect analysis of the interaction of fruit maturity degree and treatment method in affecting the seed germination potential
3 讨论
种子休眠是指胚发育完全的种子在适宜萌发的环境条件下 (水、温度、光照和空气等),在指定时间 (通常为30 d) 内不萌发或萌发率低的现象[26].从红色和紫黑色日本珊瑚树果实经去除果肉及划破内果皮处理后的种子萌发情况来看(如图1,表2所示),按照Baskin和Baskin[27]的种子休眠分类系统判断,两种成熟度果实中的种子均具有休眠,并且紫黑色果实中种子的休眠程度显著深于红色果实.换句话说,在果实进入成熟期后,随着成熟度的加深,种子休眠程度也在加深,在烟草[11]和北柴胡[28]种子的发育过程中均发现有类似现象.另外,在实验中还发现日本珊瑚树种子存在双休眠现象,在胚根突破种皮后,还要继续培养相当长的时间,胚芽才会生长,期间胚根已长得很长并生出了一些须根,按照Baskin和Baskin[26]对种子休眠的分类,日本珊瑚树种子似乎存在非深度简单上胚轴形态生理休眠,与其原变种珊瑚树的种子休眠类型相同[20-21].
红色果实的果肉和内果皮对其种子休眠与萌发均无显著影响.对于紫黑色果实而言,果肉显著抑制其种子萌发,而内果皮对其种子萌发无显著影响(如图1所示),由此也可以推论出,果实成熟度和食果鸟取食均对日本珊瑚树种子休眠与萌发具有显著影响,食果鸟对紫黑色果实的取食既促进了种子传播,也促进了种子休眠的解除和萌发;而其对红色果实的取食则仅促进了种子传播,对种子休眠和萌发无显著影响.
4 结论
日本珊瑚树种子具有非深度简单上胚轴形态生理休眠.果实进入成熟期后,随着果实颜色由红色变成紫黑色,其种子休眠程度也加深.食果鸟对紫黑色果实的取食既促进了种子传播,也促进了种子休眠的解除和萌发;而其对红色果实的取食则仅促进了种子传播,对种子休眠和萌发无显著影响.
猜你喜欢
防护林科技(2020年11期)2020-12-30 03:55:36
航天工业管理(2020年9期)2020-12-28 00:37:42
航天工业管理(2020年1期)2020-04-20 13:32:40
中国食品学报(2019年3期)2019-01-13 01:47:39
种子(2018年9期)2018-10-15 03:14:04
学苑创造·B版(2018年12期)2018-03-04 07:19:04
饮食科学(2016年10期)2016-11-19 09:16:13
蔬菜(2016年10期)2016-03-27 12:35:11
武夷学院学报(2015年3期)2015-07-18 11:03:47
作文大王·低年级(2015年8期)2015-05-30 10:48:04
