
不同弃管年限毛竹林立竹叶片功能性状的变化特征
2021-10-08 08:34何玉友陈双林郭子武汪舍平
福建农林大学学报(自然科学版) 2021年5期
何玉友, 陈双林, 郭子武, 张 玮, 汪舍平
(1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2.浙江省常山县林业技术推广站,浙江 常山 324200)
植物功能性状是植物在漫长的进化和发展过程中与外界环境相互作用的结果,能够响应生存环境的变化,并对生态系统功能产生一定的影响[1-2].受遗传与环境的双重制约,植物功能性状呈现出丰富的多样性,其变化主要表现在器官(叶片、茎、根和生殖结构)间营养元素分配及形态结构的差异上[3].叶片是植物进行光合作用和物质生产的主要器官,也是感受环境变化最敏感的器官,与植物生物量积累及其资源获取和利用能力密切相关[4-5].叶片功能性状承载较多的环境变化信息,能够直接反映植物适应环境变化所形成的生存对策,是植物功能性状的重要组成部分,具有重要的生态学意义和研究价值[6],尤其对叶片功能性状的关注越来越多[7-8].
毛竹(Phyllostachysedulis)是我国分布最广,经济、生态和社会效益俱佳的竹种,具有生长快、产量高、用途广、能持续更新、可常年收获等优良特性,是我国森林资源的重要组成部分,在竹产业中占有极其重要的地位[9-12].随着我国经济社会的不断发展,目前竹林经营环境较以往已发生很大变化.近年来,竹材价格持续走低,劳动力成本不断攀升,毛竹林经济效益降低,竹农经营积极性受挫,竹林经营强度不断下降;此外,人口城市化迁移和劳动力外流也造成农村地区劳动力短缺,导致毛竹弃管现象加剧,废弃、荒芜竹林逐年增多[13-15].浙江省是竹业强省和竹资源大省,在竹产业转型升级过程中,毛竹林弃管或失管现象也渐趋普遍[13,16-17].
由于周边各类乔、灌、草等物种渐次侵入,弃管毛竹林的物种多样性和丰富度不断提高,种间竞争逐步加剧,林分自然演替进程开启[14].在集约经营毛竹纯林向竹阔混交林直至地带性顶级群落(试验区为常绿阔叶林)演替过程中,光照、养分、水分、空间结构等毛竹生境条件及竞争关系发生剧变,势必对毛竹器官功能性状产生重要影响,进而影响到毛竹生长、发育及种群稳定,从而产生全新的生态适应策略.毛竹向外扩张已有不少研究[13,18-20],但有关竹林被阔叶树种逆向入侵方面的研究少有报道.为此,本文基于空间代替时间理论,选择不同弃管年限毛竹林为研究对象,探讨不同弃管年限毛竹林立竹叶片形态塑性、生理塑性的变化规律,旨在阐明毛竹叶片功能性状对林下植被演替过程及其环境变化的响应特征与适应策略,为开展竹林群落演替研究提供依据.
1 材料与方法
1.1 试验地概况
试验地位于浙江省常山县芳村镇棋盘山(118°40′E,29°07′N),属亚热带海洋性季风气候,气候温和湿润,雨水充沛,光照充足.年均气温17.7 ℃,绝对最低温度-11 ℃,绝对最高温度40 ℃,10 ℃以上年均积温5 468 ℃,年日照时数1 730 h,年均降雨量1 760 mm,无霜期279 d左右.试验区位于该县主要饮用水源地芙蓉水库上游,属中低山地貌,海拔450~550 m,土壤为沙质红壤.现存植被由次生常绿阔叶林、各类针(竹)阔混交林、毛竹林等类型组成,森林覆盖率75%以上.毛竹林原由村民合作组织统一经营,后因生态移民异地搬迁及环保整治,大部分毛竹纯林逐渐因弃管而荒芜,少量则由留守农户继续集约经营.竹阔混交林伴生乔木树种主要有木荷(Schimasuperba)、苦槠(Castanopsissclerophylla)、青冈(Cyclobalanopsisglauca)、红楠(Machilusthunbergii)、东南石栎(Lithocarpusharlandii)等,伴生灌木有格药柃(Euryamuricata)、赤楠(Syzygiumbuxifolium)、山矾(Symplocoscaudata)、马银花(Rhododendronovatum)、檵木(Loropetalumchinense)、胡枝子(Lespedezabicolor)等.
1.2 试验设计与样品采集
2020年4月对试验地踏查、走访,于当年7月在该地选择一片经营与弃管并存、原林分状况及经营措施相近的毛竹林作为试验区域.在试验区选择邻近立地条件基本一致而弃管年限不同的毛竹林(弃管8、20 a)及集约经营毛竹纯林(弃管0 a,对照)作为试验林,各类试验林面积不小于1 hm2.集约经营毛竹林日常主要采取季节性劈山除杂和留笋养竹、伐竹等抚育措施,不施肥;弃管毛竹林基本处于自然封育状态,人为活动干扰少或无.在每一类试验林中分别设置20 m×20 m标准样地各3个,共计9个样地.在每样地内距边缘处1 m起,沿对角线方向各平行设置3个5 m×5 m样方.测定样方内乔木、灌木的株数、株高、冠幅、胸径/地径,记录每个样地的海拔、坡度、坡位和坡向等信息.根据各样方立竹(不包括死亡和倒伏竹)调查结果,每一样地齐地伐取健康的2度标准立竹各2株,在样竹的竹冠上部、中部、下部采集竹叶混合样约150 g,放入塑料自封袋,单独编号,分别装入便携式冰盒中,带回实验室冷藏.样地基本情况见表1、2.
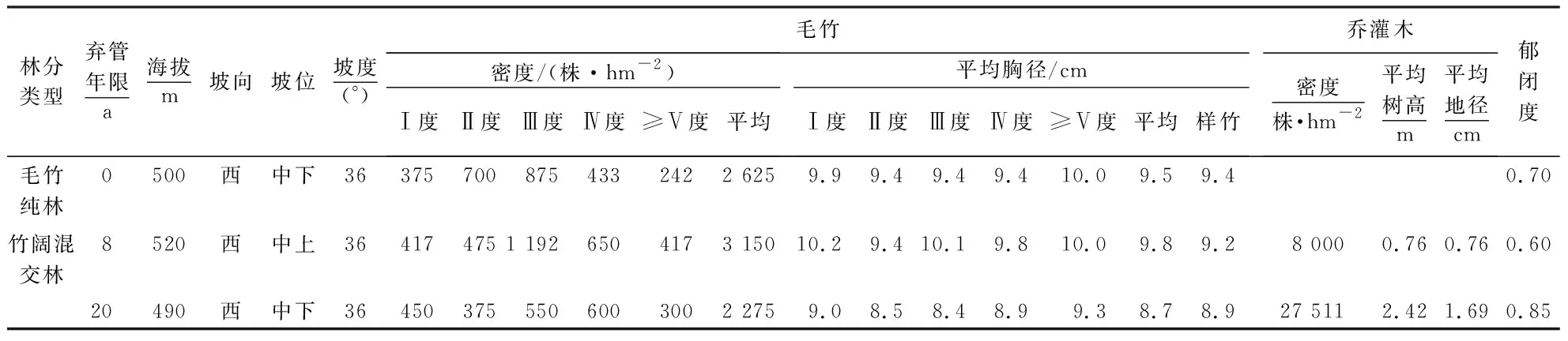
表1 不同弃管年限毛竹林的基本情况Table 1 Basic information on P.edulis stands unattended for different years

表2 不同弃管年限毛竹林的土壤酸碱度及养分含量1)Table 2 Soil pH and nutrient contents of P.edulis stands unattended for different years
1.3 测定方法
1.3.1 叶片性状测定 每株样竹随机选取10片生长正常、叶形完整、无病虫害的成熟叶片[9],用湿润纸巾反复擦净正反叶面,待叶面风干后测定叶形指标.选用精度0.01 mm数显游标卡尺,在叶片中部沿主脉一侧测量叶片厚度.用Epson Perfection V330 Photo扫描仪扫描叶片,用万深LA-S系列植物图像分析系统对扫描图进行分析处理,得到叶长、叶宽、叶周长、叶面积、叶长宽比等指标.
将叶片放入烘箱103 ℃杀青30 min,然后85 ℃烘干至恒重,称量得叶干重.
比叶面积由叶面积与叶干重相除而得;叶组织密度为叶干重与叶体积之比,即单位体积的叶片干重.
1.3.2 色素含量测定 选取健康、正常叶片3~5片,擦净叶表污染物,再将被测叶片剪成宽度小于1 mm的细丝,然后充分剪碎、混匀.在50 mL锥形瓶中加入体积比为1∶1的丙酮与无水乙醇混合液5 mL,用0.000 1 g电子天平称取碎叶样品约0.05 g,放入瓶中,于室内避光处浸泡提取24 h;提取液过滤后,取1 mL提取液,加入丙酮与无水乙醇各1 mL,混合均匀.以丙酮与无水乙醇(体积比为1∶1)混合液作为空白对照,分别在波长645、663和470 nm处测定光密度,然后根据文献[21]分别计算出叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素[Chl(a+b)]、类胡萝卜素(Car)含量.依同法制备样品,参照文献[22]测定竹叶花青素(Ant)含量.
1.4 数据处理
试验数据均表示为平均值±标准差.采用单因素方差分析比较某一测定指标的差异显著性,多重比较采用LSD法,显著水平(α)=0.05.用主成分分析法筛选叶功能性状变化的主要指标,性状间相关性采用Pearson双侧检验.所有数据用SPSS 26.0软件分析,并用Excel 2007作图.
2 结果与分析
2.1 不同弃管年限毛竹林叶片形态性状的变化
随着毛竹林弃管年限的延长,叶长、叶宽、叶周长、叶面积、叶干重及比叶面积总体呈下降趋势,降幅为6.5%~18.8%,以叶面积降幅最大,但各性状在不同弃管年限竹林间差异不显著(P>0.05).而叶组织密度则总体上升高,增幅约5.1%,在不同弃管年限间差异也不显著(P>0.05).叶长/叶宽、叶片厚度呈倒“V”形变化,与弃管8 a竹林相比,变幅为1.3%~7.1%,不同弃管年限间也无显著差异(P>0.05).各叶形性状变异系数较小,为3.93%~14.13%,其中以叶面积、叶干重两性状变异稍大(表3).由上可知,弃管竹林在自然演替进程中,毛竹叶形性状随生境的改变出现一定程度的适应性变化.

表3 不同弃管年限毛竹林立竹叶片的形态性状1)Table 3 Leaf morphological characteristics of P.edulis stands unattended for different years
2.2 不同弃管年限毛竹林叶片光合色素的变化
图1显示,随着毛竹林弃管年限的延长,竹叶光合色素含量先降后升;集约经营(弃管0 a)与弃管8 a竹林相比,Chla、Chlb、Chl(a+b)和Car存在显著差异,增幅分别为50.6%、65.1%、53.9%、30.0%;弃管20 a竹林与弃管8 a竹林相比,Chla、Chlb、Chl(a+b)和Car存在显著差异,增幅分别为60.4%、72.9%、63.2%、37.1%.集约经营(0 a)与弃管20 a竹林间无显著差异.各光合色素含量表现为:弃管20 a竹林≥集约经营(0 a)竹林>弃管8 a竹林.Ant含量随弃管年限的延长而下降,降幅约为22.4%,不同弃管年限竹林间无显著差异.由上可知,毛竹叶片色素含量随弃管年限的变化与色素种类有关,光合色素含量变化显著且呈上升趋势,而Ant呈下降趋势,变化不显著.
从图2可知,Chl(a+b)/Car、Chl(a+b)/Ant、Car/Ant随竹林弃管年限的延长呈先降后升的“V”形变化趋势,与所涉光合色素的变化趋势基本一致,而Chla/Chlb呈倒“V”变化.Chl(a+b)/Car在集约经营(0 a)及弃管20 a竹林间无显著差异,但与弃管8 a竹林差异显著; Chla/Chlb、Chl(a+b)/Ant、Car/Ant在不同弃管年限竹林间差异不显著.Chl(a+b)/Ant变异最大,变异系数达40.4%;Chla/Chlb变异系数最小,为5.3%.Chla/Chlb大小表现为:弃管8 a竹林>弃管20 a竹林>集约经营(0 a)竹林.其它色素比值则表现为:弃管20 a竹林>集约经营(0 a)竹林>弃管8 a竹林,与光合色素含量高低序次一致.
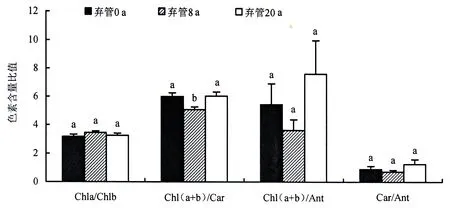
图2 不同弃管年限毛竹林立竹叶片色素含量的比值Fig.2 Ratios of different photosynthetic pigments in leaves of P.edulis stands unattended for different years
2.3 不同弃管年限毛竹林叶片功能性状之间的关系
相关分析(表4)表明,不同弃管年限毛竹林立竹叶片光合色素Chla、Chlb、Chl(a+b)、Car间呈显著或极显著正相关,说明光合色素含量变化具有较好的协同性;Ant与各叶形性状及光合色素各组分间均无显著相关关系.

表4 不同弃管年限毛竹林叶片功能性状之间的相关性1)Table 4 Correlation between leaf functional traits of P.edulis stands unattended for different years
不同弃管年限毛竹林叶形性状与光合色素含量间的相关性差异较大.叶面积与叶干重呈正相关关系,集约经营竹林及弃管8 a竹林的叶面积与叶干重显著相关.比叶面积与叶片厚度、叶组织密度、叶干重呈负相关关系,其中集约经营和弃管20 a竹林的叶片厚度、叶组织密度及弃管8 a竹林的叶干重呈显著负相关.光合色素Chla、Chlb、Chl(a+b)、Car与各叶形性状间的相关性基本一致,如与叶面积、比叶面积等面积类性状呈正相关,与叶组织密度、叶干重、叶片厚度等质量结构属性呈负相关,但相关程度各异.仅弃管20 a竹林中的光合色素与比叶面积、叶组织密度及弃管8 a竹林的Car与叶片厚度之间的相关性达显著或极显著程度.总体而言,随着弃管年限的延长和光合色素含量的提高,毛竹林叶片功能性状间的相关关系渐趋增强.
2.4 不同弃管年限毛竹林叶片功能性状的主成分分析
从表5可知,前3个主成分的贡献率分别为53.95%、20.41%和15.07%,三者累积贡献率达89.43%,可以反映不同弃管年限毛竹林诸多叶片功能性状的大部分变异信息.在第1主成分中,Chla、Chlb、Chl(a+b)和Car具有较高载荷,反映了叶片光合色素含量的信息.叶面积在第2主成分中具有较高载荷,主要代表了叶面积塑性指标信息.第3主成分表达的是以比叶面积、叶干重、叶组织密度等有关叶片质地、结构为主的综合信息.据上分析可知,光合色素含量是反映弃管竹林叶功能性状变化特征的主要指标.

表5 不同弃管年限毛竹林叶片功能性状的主成分分析Table 5 Principal component analysis of leaf functional traits of P.edulis stands unattended for different years
3 结论与讨论
3.1 不同弃管年限毛竹林叶片形态性状的变化特征
叶功能性状随着环境变化具有较强的可塑性和环境敏感性[23-24].本研究结果显示,随着弃管年限的延长,毛竹林立竹叶片的叶长、叶宽、叶面积、叶干重、叶周长及比叶面积呈减小的变化趋势;叶组织密度则呈增大的变化趋势;而叶长/叶宽、叶片厚度呈倒“V”形的变化趋势.但叶片形态性状总体上未达显著差异,与刘希珍等[18]的研究结果部分一致.集约经营毛竹林的年龄结构、空间分布、径级大小等较均匀合理,故叶面积、比叶面积等增大,叶长/叶宽、叶片厚度、叶组织密度等变小,以适应资源相对富裕的环境,提高光合生产能力[5,25-27],所以林分生产力水平较高,生态稳定性好.本研究结果表明弃管8 a毛竹林由于植被低矮稀疏、盖度低,林分比较通透,水肥等立地条件相对较差,且立竹密度较大,故叶片厚度、叶长/叶宽增大以降低蒸腾作用,增加组织抗性和逆境适应能力[28];比叶面积变小以适应资源贫瘠和干旱的环境[5,25-26].竹林弃管20 a后,大量壳斗科、山茶科等乔灌树种入侵并发育充分,林分郁闭度大幅提高,群落结构更趋复杂[29].本研究结果表明林地枯枝落叶层增厚,土壤养分、有机质含量提高,酸性渐减,土壤理化性质得到较大改善[30-31],所以弃管20 a毛竹林的叶长/叶宽、叶片厚度和集约经营竹林相似.弃管20 a后竹林土壤状况更利于出笋长竹,然而毛竹林的径级下降[14],密度减小,优势度降低,因种间竞争加剧和环境资源制约,以及竹林结构和功能退化之故,致使竹叶叶面积、比叶面积、叶干重等相应变小,叶组织密度增大.这有利于叶片养分积累和营养保存,增强其耐受性和防御能力[28],与毛竹向周边林分扩张的适应性变化相似[18].
叶形态性状变化受物种遗传特性和环境因子的共同影响,而物种遗传特性占有绝对的解释力[32].本研究结果也显示,弃管竹林随弃管年限的延长,毛竹各叶形性状未因生境改变而产生显著差异,但其对环境变化的响应策略和变化趋势与众多研究结果基本相符.刘希珍等[18]研究显示叶干物质含量在毛竹不同扩张阶段差异显著,可能是由于演替方向不同,扩张毛竹所处生境相异.本研究中各弃管竹林毛竹始终居林冠上层,而伴生乔灌木尚处中幼龄和林下受压状态,林分状况两者迥异.
3.2 不同弃管年限毛竹林叶片光合色素的变化特征
本试验结果显示,光合色素含量随竹林弃管年限的延长先降后升,在弃管20 a竹林中最高,在弃管8 a竹林中最低,不同弃管年限间差异显著,说明光合色素对环境变化敏感,对环境变化快速适应.Ant含量和Chla/Chlb、Chl(a+b)/Ant、Car/Ant差异不显著,表明对环境变化的敏感性差,这与大部分研究结果[33]基本一致.研究[34-35]表明,当土壤存在氮元素缺乏、干旱等胁迫时,叶绿素含量会随之下降.
植物的叶色和光合性能不仅受叶片色素含量绝对值的影响,更与色素含量间的比值有关[36].Chla/Chlb是衡量植物耐荫性的重要指标,一般阳生植物与阴生植物各为3.0、2.3[37].本研究试验林中,Chla/Chlb为3.1~3.4,以弃管8 a竹林最高.研究[38]表明,光照、温度、水分等对Chla/Chlb影响较大.弃管8 a竹林可能是因干燥强光生境造成Chlb含量降低,从而提高了Chla与Chlb的比值.
3.3 不同弃管年限毛竹林叶片功能性状的相关性分析
研究[3,18,39-40]表明:比叶面积与叶组织密度、叶片厚度、叶干重呈负相关关系,叶面积与叶干重呈正相关;光合色素各组分Chla、Chlb、Chl(a+b)和Car显著正相关[41-42],反映出性状间关联关系的共性与普遍性[43].本文研究结果与上述结论基本一致,但在关联程度及部分性状间的相关关系上仍有差别,这是由毛竹特有的遗传特性或群落环境不同造成的.本研究也发现,毛竹叶形性状与光合色素各组分间的相关性随弃管年限的延长总体上有增强的趋势,并且光合色素可作为反映叶功能组合性状变化的主要指标.
猜你喜欢
农学学报(2022年8期)2022-11-16
农业与技术(2022年16期)2022-09-01
热带农业工程(2021年5期)2021-12-06
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
绿色科技(2017年15期)2017-09-01
绿色科技(2017年9期)2017-06-01
农家科技下旬刊(2017年2期)2017-03-27
