
盐胁迫对水稻颖花形成及籽粒充实的影响
2021-10-05 13:51:32韦还和张徐彬葛佳琳孟天瑶陈英龙戴其根
作物学报 2021年12期
韦还和 张徐彬 葛佳琳 陈 熙 孟天瑶 杨 洋 熊 飞 陈英龙 戴其根,,*
1 江苏省作物遗传生理重点实验室 / 江苏省作物栽培生理重点实验室 / 江苏省粮食作物现代产业技术协同创新中心 / 扬州大学水稻产业工程技术研究院, 江苏扬州 225009; 2 扬州大学教育部农业与农产品安全国际合作联合实验室 / 扬州大学农业科技发展研究院, 江苏扬州225009; 3 扬州大学生物科学与技术学院, 江苏扬州 225009
我国有234万公顷沿海滩涂和1亿公顷内陆盐碱地, 是我国最重要的后备耕地资源[1]。水稻常作为盐土改良的重要粮食作物, 有关盐胁迫下水稻生长发育与产量形成特征一直备受关注[2-3]。研究表明,盐胁迫对水稻植株生长发育的伤害主要包括渗透胁迫和离子胁迫。渗透胁迫主要是由于土壤中高浓度的盐分降低了土壤水势, 使植株吸水困难, 抑制植株生长; 离子胁迫则是由于植株叶片中Na+的过量积累常导致K+、Ca2+和磷素吸收受抑, 从而产生单盐毒害[4-5]。盐胁迫下, 水稻植株光合作用受到抑制,不仅加剧了光合产物在主茎与分蘖之间的竞争, 引起部分分蘖困难且也难以获得充足的养分而未能成穗; 而且加快了花后植株衰老进程, 导致群体库容充实差、产量低[6-8]。当前, 我国正大力开发利用沿海滩涂和内陆盐碱地发展水稻生产[9-10]。但总体上,沿海滩涂和内陆盐碱地种植的水稻产量偏低且稳产性较差[11-12]。每穗颖花数是决定水稻丰产性与稳产性的重要因子, 取决于颖花的分化与退化2个过程。水稻穗部颖花分化和退化, 既受植株营养物质供应的多寡、碳/氮比值的平衡以及幼穗器官生理活性高低等诸多内在生理因素的影响[13-15], 也易受外在环境条件和栽培措施的影响[16-18]。环境条件方面, 王亚梁等[16]研究表明, 水稻穗分化期高温胁迫会加剧颖花退化, 造成每穗颖花量显著下降; 抑制花药发育, 降低花粉活力, 导致籽粒充实性状差。栽培措施方面, 陈培峰等[17]研究表明, 与浅水灌溉相比, 轻干湿交替灌溉利于促进稻穗枝粳和颖花的分化, 尤其是促进稻穗中下部二次颖花的现存数的增加, 显著提高了中、弱势粒的粒重和充实度。目前, 关于盐胁迫下水稻穗部颖花分化与退化及籽粒充实的报道较少, 且缺少系统研究。为此, 本研究以江苏沿海滩涂大面积种植的常规粳稻南粳9108和盐稻12为试材, 研究分析盐胁迫对稻穗颖花形成、花器官发育及籽粒充实的影响, 旨在为沿海滩涂水稻高产栽培与调控措施提供理论与实践支撑。
1 材料与方法
1.1 供试材料
以江苏沿海滩涂大面积种植的常规粳稻南粳9108和盐稻12为供试材料。
1.2 试验设计与栽培管理
2019—2020年, 在扬州大学农学院试验农场进行盆栽试验, 盆钵直径25 cm、高30 cm。盆钵所装土壤取自试验农场稻田耕层, 土壤类型为沙壤土,20 cm土壤耕层中含全氮0.13%、碱解氮89.4 mg kg–1、速效磷32.9 mg kg–1、速效钾80.1 mg kg–1, 每盆装土18 kg。
设置对照(check, CK, 盐浓度0%)、中盐(medium-salinity stress, MS, 盐浓度0.15%)和高盐(high-salinity stress, HS, 盐浓度0.3%)共3个盐浓度梯度处理, 每处理50盆。根据试验设计的含盐量梯度和盆钵土的重量来计算NaCl海盐的重量, 移栽前3 d将对应重量的Q/ZLY型速溶海水晶(浙江蓝海星盐制品厂生产, 天然海水晒制, 基本保留了海水的主要成分)均匀撒施在对应处理盆钵中, 并与土壤充分搅拌均匀。盆栽场上方建有遮雨棚, 以防因降雨引起池内盐分变化。2年中, 南粳9108和盐稻12均于5月20日播种, 采用塑料软盘旱育秧, 秧龄25 d后移至盆钵, 每盆4穴, 每穴4株苗。每盆基施2 g尿素和0.5 g磷酸二氢钾, 移栽后7 d每盆施0.5 g尿素, 穗分化期每盆施0.6 g尿素。移栽后, 各盆栽处理建立1~2 cm浅水层, 以湿润灌溉为主; 穗分化阶段与抽穗扬花期始终保持2~3 cm水层; 灌浆结实期实行干湿交替, 自1~2 cm浅水层自然落干至土壤水势–15 kPa, 然后灌1~2 cm浅水层, 再落干, 如此循环,直至成熟。盆钵内安装真空表式负压计(中国科学院南京土壤研究所生产)监测土壤水势。按常规高产栽培管理病虫草害。
1.3 测定项目与方法
1.3.1 主茎穗颖花分化与退化及颖花大小 于抽穗期, 集中取样1次, 各处理选取大小一致主茎穗20个。以单茎为单位, 观察每穗一次和二次枝粳上的颖花现存数与退化数, 退化的颖花为具有白色的小颖花状薄膜痕迹, 部分可用肉眼观察计数, 部分颖花退化数痕迹较小在体视显微镜(OLYMPUS,SZX16)下观察计数。借助万深考种仪(SC-G)测量主茎穗颖花大小。
颖花现存(退化)数 = 一次枝粳颖花现存(退化)数 + 二次枝粳颖花现存(退化)数;
颖花分化数 = 颖花现存数 + 颖花退化数;
颖花退化率(%) = (颖花退化数 / 颖花分化数)×100;
一次(二次)枝粳颖花分化数 = 一次(二次)枝粳颖花现存数 + 一次(二次)枝粳颖花退化数;
一次(二次)枝粳颖花退化率 = 一次(二次)枝粳颖花退化数×100 / 一次(二次)枝粳颖花分化数。
1.3.2 主茎穗花药形态 于主茎稻穗抽出约20%~30%时集中取样1次。各处理选取当天开放的花药30个, 在体视显微镜下(OLYMPUS, SZX16)观察花药形态, 并照相。之后, 利用测微尺测量花药长度(length, L)和宽度(width, W)。花药体积(V)按公式V = 0.34LW2计算[19]。
1.3.3 主茎穗花药开裂率 取样方法与时期同1.3.2。各处理选取当天开放的花药30个, 在体视显微镜下(OLYMPUS, SZX16)观察花药开裂情况, 计算花药开裂率。
花药开裂率(%) = (完全开裂花药数+部分开裂花药数) / 镜检花药总数×100。
1.3.4 主茎穗花粉活力 取样方法与时期同1.3.2。各处理选取当天开放的颖花3枚, 用镊子取下每枚颖花上的全部花药挤碎, 用1% I2-KI溶液染色并于生物显微镜下(OLYMPUS, BX61)选取3个视野统计可染率, 能染色的花粉即具有活力的花粉。
1.3.5 主茎穗穗部性状与实产 于成熟期, 各处理选取大小一致主茎穗40个, 测定单穗重、每穗粒数、结实率和粒重。各处理选取植株长势基本均匀,且未经取样测定的10盆稻株, 每盆单独手工脱粒计产。
1.4 数据处理
运用Microsoft Excel软件录入数据、计算, DPS软件作统计分析, SigmaPlot软件作图。
2 结果与分析
2.1 产量与主茎穗部性状
与对照相比, 盐胁迫显著降低了南粳9108和盐稻12产量。中盐胁迫下, 南粳9108和盐稻12产量降幅分别为18.7%和24.3%; 高盐胁迫下, 南粳9108和盐稻12产量降幅分别为54.5%和58.6% (表1)。盐胁迫处理下水稻主茎单穗重和每穗粒数显著低于对照。2年中, 中盐和高盐胁迫下南粳9108每穗粒数较对照分别低8.6%和19.8%, 盐稻12较对照分别低8.0%和25.9%; 穗部籽粒充实性状方面, 盐胁迫处理显著降低了南粳9108和盐稻12穗部籽粒的结实率和粒重, 2年中, 中盐和高盐胁迫下盐稻12穗部粒重分别为24.4 mg和22.9 mg, 较对照分别低1.2 mg和2.7 mg (表2)。

表1 盐胁迫对水稻产量的影响Table 1 Effects of salinity stress on grain yield in rice
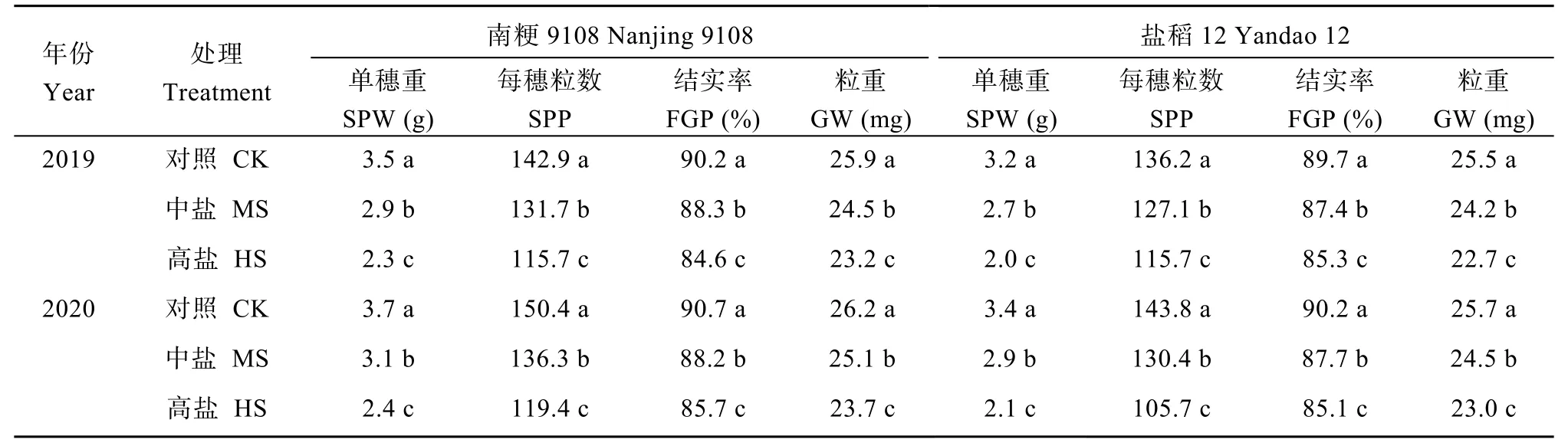
表2 盐胁迫对水稻主茎穗部性状的影响Table 2 Effects of salinity stress on panicle traits of the main stem in rice
2.2 主茎穗部颖花分化与退化
与对照相比, 盐胁迫处理降低了水稻主茎颖花分化数和颖花现存数。中盐胁迫下南粳9108和盐稻12主茎颖花分化数较对照分别降低了4.4%和3.5%,高盐处理下则分别降低了12.1%和11.2%。盐胁迫处理显著提高了主茎颖花退化数与退化率; 2年中, 中盐和高盐胁迫下南粳9108主茎颖花退化数分别为15.2和22.3, 较对照分别增加了72.7%和153.4% (表3)。对南粳9108和盐稻12而言, 中盐胁迫下一次枝粳上颖花分化数和颖花现存数均与对照差异不显著,高盐胁迫下一次枝粳上颖花分化数和颖花现存数则显著低于对照。与对照相比, 中盐和高盐胁迫处理提高了主茎一次枝粳颖花退化数与退化率(表4)。中盐和高盐胁迫下, 主茎二次枝粳颖花分化数与颖花现存数均显著低于对照, 颖花退化数与颖花退化率则显著高于对照。中盐和高盐胁迫下, 南粳9108主茎二次枝粳颖花退化数较对照分别增加了52.7%和112.7%, 盐稻12主茎二次枝粳颖花退化率则较对照分别增加了51.7%和108.8% (表5)。

表3 盐胁迫对水稻主茎穗部颖花分化与退化的影响Table 3 Effects of salinity stress on differentiated and retrograded spikelets of the main stem in rice

表4 盐胁迫对水稻主茎一次枝粳颖花分化与退化的影响Table 4 Effects of salinity stress on differentiated and retrograded spikelets of the primary branches on the main stem of rice

表5 盐胁迫对水稻主茎二次枝粳颖花分化与退化的影响Table 5 Effects of salinity stress on differentiated and retrograded spikelets of the secondary branches on the main stem in rice
2.3 主茎花药性状、花粉活力与颖花大小
与对照相比, 盐胁迫处理降低了水稻主茎花药长度、宽度与体积。中盐和高盐胁迫下, 南粳9108花药长度2年平均为1.71 mm和1.60 mm, 较对照分别下降了4.7%和11.1%; 盐稻12花药长度2年平均为1.80 mm和1.66 mm, 较对照分别下降了6.5%和13.8% (表6)。中盐和高盐胁迫下南粳9108花药开裂率分别为69.9%和55.6%, 盐稻12则分别为68.0%和54.0%, 均显著低于对照(图1)。对于南粳9108和盐稻12而言, 盐胁迫处理均显著降低了花粉活力。中盐和高盐胁迫下南粳9108花粉活力分别为81.6%和64.7%, 较对照分别降低了13.6%和31.5% (图2)。与对照相比, 盐胁迫处理降低了水稻主茎穗部颖花的长度和宽度。以南粳9108为例, 高盐胁迫下主茎穗部颖花长度和宽度分别为6.63 mm和3.49 mm, 较对照处理分别低6.5%和6.1% (表7)。
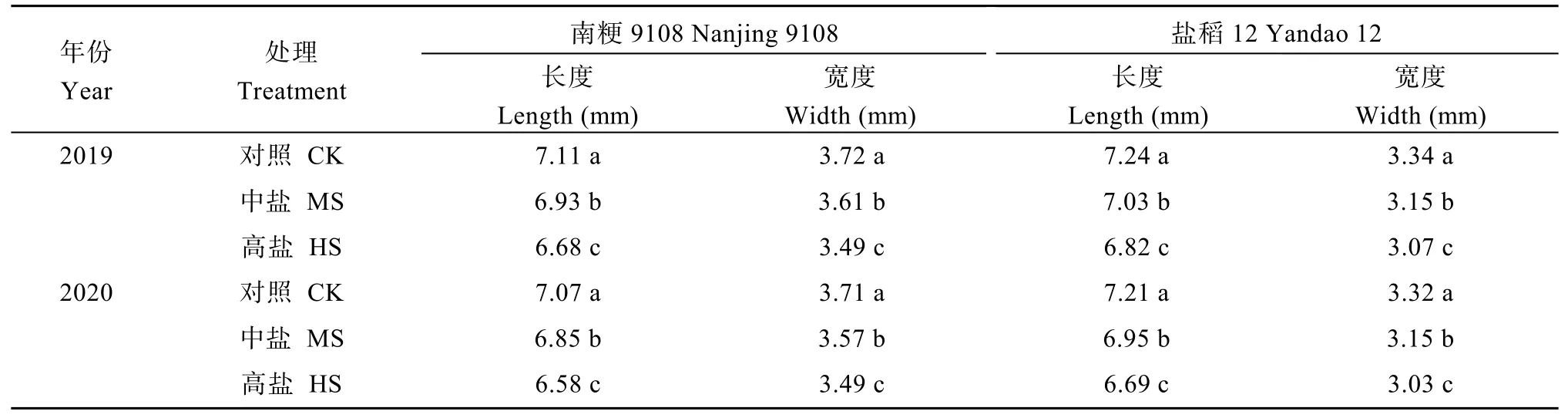
表7 盐胁迫对水稻主茎穗颖花大小的影响Table 7 Effects of salinity stress on size of spikelets on the main stem in rice
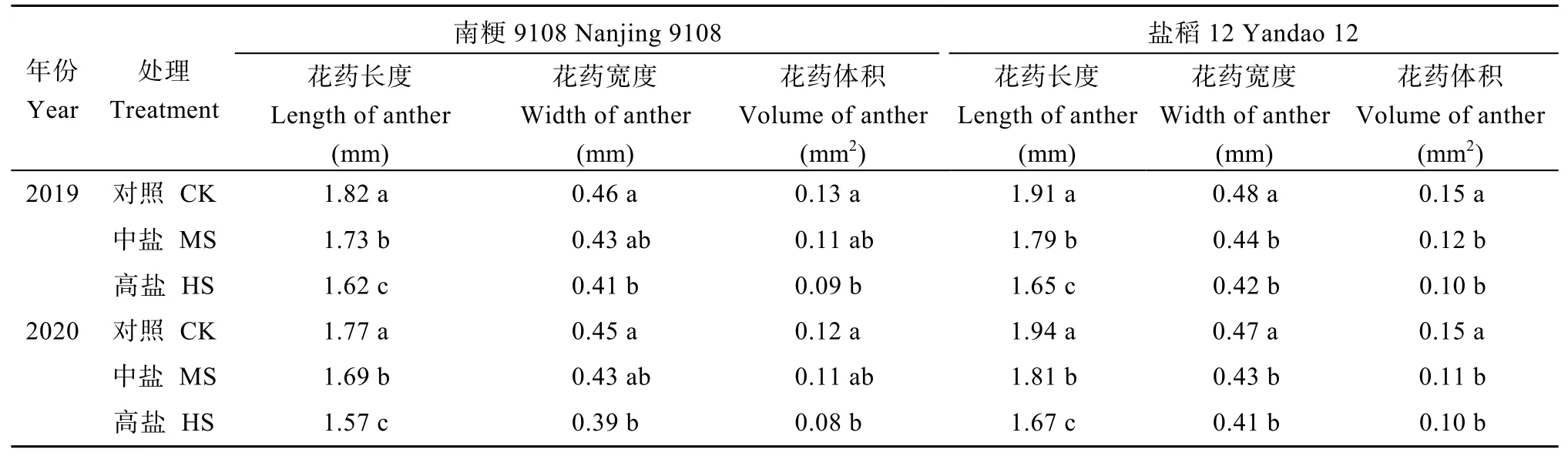
表6 盐胁迫对水稻主茎花药大小的影响Table 6 Effects of salinity stress on size of anther of the main stem in rice
2.4 相关性分析
相关性分析结果表明, 颖花长度与产量、每穗粒数、结实率和粒重呈极显著正相关。一次和二次枝粳颖花分化数和现存数与产量、每穗粒数、结实率和粒重呈极显著正相关, 颖花退化数和颖花退化率则与产量、每穗粒数、结实率和粒重呈极显著负相关。此外, 花药长度与宽度、花药开裂率、花粉活力与产量、每穗粒数、结实率和粒重均呈显著或极显著正相关(表8)。

表8 盐胁迫处理下水稻穗部性状的相关性分析Table 8 Correlation analysis of panicle traits under salinity stress in rice
3 讨论
本试验条件下, 南粳9108和盐稻12在中盐和高盐胁迫下产量降幅分别为1 8.7%~2 4.3%和54.5%~58.6% (表1), 表明盐胁迫显著降低了水稻产量, 这与此前的研究结果基本一致[6,8,20]。有关盐胁迫对水稻产量构成因素的影响已有较多研究报道,但研究结果并不完全一致。周根友等[6]、孙现军等[21]研究表明, 与非盐胁迫相比, 盐胁迫导致水稻产量显著下降的主要原因是每穗粒数和粒重的降低; 而胡博文等[22]、荆培培等[23]研究表明, 盐胁迫下水稻群体穗数、每穗粒数、结实率和粒重均降低, 从而使水稻产量显著下降。本试验条件下, 与对照相比,中盐和高盐胁迫均显著降低了南粳9108和盐稻12的每穗粒数、结实率和粒重(表2); 此外, 笔者此前的研究表明[8], 盐胁迫下水稻分蘖发生率与成穗率均较低, 单株与群体成穗数少。因此, 库容量偏小与库容充实差共同制约了盐胁迫下水稻产量的提升,这与胡博文等[22]、荆培培等[23]的研究结果基本一致。
目前有关盐胁迫影响水稻颖花形成的研究报道较少。本研究结果表明, 与对照相比, 盐胁迫处理降低了水稻颖花分化数, 提高了颖花退化数, 每穗颖花数显著下降; 从影响过程来看, 盐胁迫下南粳9108和盐稻12颖花退化数的增加幅度高于颖花分化数的下降幅度, 因此, 盐胁迫下颖花退化数的增加是导致每穗颖花数下降的关键因素(表2和表3)。盐胁迫导致水稻穗部颖花数增加的原因可能有以下2点: (1) 植株内源激素比例失衡。水稻植株内源激素平衡程度是影响颖花退化的关键因素之一。水稻穗分化期尤其是减数分裂期较低的内源油菜素甾醇(BRs)和多胺(PAs)水平以及较高的乙烯水平与颖花退化密切相关, 提高BRs、PAs水平或PAs与乙烯的比值可有效减少颖花退化[24-25]。此前研究表明[26-27],盐胁迫下水稻植株乙烯释放量急剧上升, PAs水平和PAs与乙烯的比值则显著下降, 加剧了穗部颖花的退化。(2) 盐胁迫易造成植株能荷水平过低及活性氧过度积累, 使得膜脂过氧化伤害和小穗程序性细胞死亡, 导致颖花退化数的增加[28-29]。
关于不同枝粳上颖花分化与退化变化的比较已有相关研究报道[15,18,30]。董明辉等[18]研究表明, 麦秸还田与施氮量处理均对枝梗与颖花分化数、退化数/率、现存数产生显著影响, 对二次枝梗和二次颖花的影响大于一次枝梗和一次颖花。王亚梁等[30]研究表明, 穗分化期高温胁迫抑制水稻颖花分化, 加剧颖花退化; 高温对二次枝粳上颖花分化与退化的影响大于一次枝粳。本试验条件下, 盐胁迫处理降低了一次和二次枝粳颖花分化数和现存数, 增加了一次和二次枝粳颖花退化数; 盐胁迫下二次枝粳颖花分化数和现存数的下降幅度和颖花退化数的增加幅度均高于一次枝粳(表4和表5), 表明盐胁迫对一次和二次枝粳颖花形成的影响存在差异, 以对二次枝粳颖花形成的影响更大。
本研究表明, 与对照相比, 盐胁迫处理下水稻籽粒充实性状(结实率和粒重)显著变劣(表2)。水稻花器官正常发育是籽粒充实饱满以及获取高产的关键因素之一。花器官的发育极易受到环境胁迫因子的影响[14-15,19,31]。温度胁迫方面, 张荣萍等[19]研究表明, 开花期低温胁迫影响水稻结实率的主要原因是低温胁迫影响花药发育成熟, 使花药不能正常开裂、散粉不足, 可育花粉率和柱头花粉萌发率降低,直接影响了受精结实。水稻抽穗扬花期遇到高温胁迫, 对水稻的花粉粒直径、花粉活力和花药开裂率等均会产生不同程度的影响[14,31]。本试验条件下, 与对照相比, 盐胁迫降低了花药大小及其开裂率与花粉活力(表6, 图1, 图2), 表明盐胁迫下水稻花器官受到伤害, 使花药开裂受阻, 导致散发到柱头上的花粉量不足, 进而影响授粉受精, 引起结实率下降(表8)。水稻粒重取决于颖花大小和籽粒饱满度, 前者决定了库容潜力, 后者跟同化物的供应有关[32-33]。本研究结果表明, 与对照相比, 盐胁迫处理降低了水稻颖花长度和宽度(表7), 抑制了库容潜力的提升;此外, 盐胁迫下植物光合作用受到抑制, 植株衰老进程加快, 花后干物质积累量少, 导致籽粒充实饱满度差[8,34]。因此, 盐胁迫抑制了水稻库容和籽粒充实饱满度的提升, 从而显著降低了粒重(表2)。
本研究在盆栽条件下, 模拟研究了不同盐胁迫浓度下水稻颖花分化与退化的变化趋势及花器官发育特点, 明确了盐胁迫对水稻颖花形成及籽粒充实的影响, 研究结果对于沿海滩涂水稻高产栽培与调控措施具有一定指导意义。考虑到盆栽条件与大田实际条件的差异, 本研究中的部分研究结果仍需进一步在沿海滩涂田间试验中加以验证。
4 结论
盐胁迫对水稻颖花形成及籽粒充实有显著影响。与对照相比, 盐胁迫抑制水稻颖花分化, 促进颖花退化, 每穗颖花数显著下降。影响花器官正常发育, 花药大小、花药开裂率和花粉活力下降; 缩短颖花长度和宽度, 降低籽粒饱满度, 使籽粒充实性状显著变劣。
猜你喜欢
Acta Mathematica Scientia(English Series)(2024年1期)2024-03-23 08:02:54
江西农业大学学报(2023年2期)2023-05-08 01:32:10
作物杂志(2022年6期)2022-02-03 04:56:22
北方水稻(2021年6期)2021-02-17 07:17:50
中国盐业(2018年24期)2018-04-04 05:33:40
中国盐业(2018年24期)2018-04-04 05:33:32
中国盐业(2018年23期)2018-03-30 01:29:22
中国盐业(2018年21期)2018-03-05 08:06:06
广西植物(2016年10期)2016-11-11 06:51:39
西南农业学报(2016年4期)2016-05-17 05:41:45
