
鲐鱼生活史及模型应用研究进展
2021-09-17 09:55李曰嵩邢宇娜潘灵芝张瑜余为
大连海洋大学学报 2021年4期
李曰嵩,邢宇娜,潘灵芝,张瑜,余为
(1.上海海洋大学 海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发教育部重点实验室,上海 201306;4.远洋渔业协同创新中心,上海 201306;5.国家海洋局东海预报中心,上海 200081)
鱼类资源是一种可自我更新的持续性资源,资源的动态和变化并非完全受捕捞的影响,其更新能力也取决于种群的自我调节能力,即取决于其世代的强弱[1]。渔业资源的补充量主要取决于鱼类早期的存活率,一旦海洋环境条件发生了微小的改变,就会对生活史中最脆弱阶段的鱼卵和仔幼鱼生长、发育、存活产生较大影响。鲐鱼Scomberjaponicus为中上层鱼类,其生态进化方向为r选择型,所以鲐鱼世代的更替速度较快。若鲐鱼资源受到破坏,只要能够及时进行科学管理并采取必要措施,其资源量就会得到较快恢复,故鲐鱼资源可持续开发利用具有较广阔的前景。若能找到鱼类生活史中的影响因素,探明导致鱼类资源量发生变化的动力学原因,就可以更好地进行渔业资源的保护、管理、利用和开发。如今全球气候发生变化,海平面和温度等外界因素的改变必定会对鱼类产生一些影响,因此,及时了解环境变化对鲐鱼的影响并利用模型等工具进行模拟预测显得十分重要,对更加合理利用鲐鱼资源具有重要的意义。
鲐鱼属于大洋暖水性中上层鱼类,是太平洋、大西洋、印度洋的主要经济鱼类之一,其中,亚太海域主要分布于东海、黄海、日本海及西北太平洋海域[2],最北可达鄂霍次克海,涉及中国、朝鲜、韩国、日本及俄罗斯远东等国家海域(图1、图2)。国内渔场主要有海洋岛、连青石、钓鱼岛、大沙及沙外等,国外渔场有北海道渔场、北海渔场、秘鲁渔场等。鲐鱼是一种重要的渔业资源,尤其是在围网渔业中,中国、日本、韩国和一些其他国家都对其进行了较好的开发利用。有研究认为,目前鲐鱼资源出现了下降趋势[3-5],因此,对鲐鱼的保护和可持续发展利用非常重要。本研究中,综述了国内外学者对鲐鱼的生物学特征、早期生活史概况及模型应用研究和相关成果,并针对目前研究中存在的问题提出了今后重点研究的方向,旨在为鲐鱼资源评估和资源补充量预测提供科学依据。
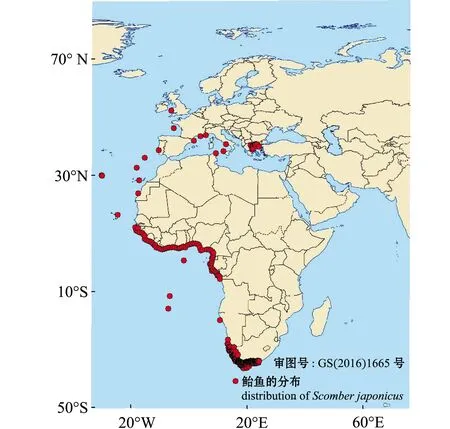
图1 鲐鱼在大西洋的分布

图2 鲐鱼在太平洋的分布
1 鲐鱼的基本生物学特征
鲐鱼又称日本鲭,属硬骨鱼纲Steichthyes鲈形目Perciformes鲭亚目Scombroidei鲭科Scombridae鲐属Scomber,体型呈纺锤状,背部为淡绿色,腹部为银灰色,有蓝黑色斑点[6],如图3所示。鲐鱼有青花鱼、鲭鱼、花池鱼、油胴鱼、花巴、花鳀等别称。在垂直分布上,鲐鱼在春、夏季时一般栖息于海水中上层,在温跃层之上的区域活动,到了繁殖季节一般会聚集为大群到水面活动,具有趋光繁殖的特性。中国近海鲐鱼索饵期间一般活动在20~50 m水层,而到了冬季则下移至100~150 m深处越冬。栖息水温一般为10~27 ℃,最适栖息水温为25 ℃左右[7],产卵场水温一般为15~20 ℃[8]。

图3 日本鲐 [9]
1.1 产卵
鲐鱼分批产卵,生殖季节中至少排卵3次,放卵时间集中在夜里到黎明前后,或是从傍晚到半夜。怀卵量为25~263万枚。卵浮性,形态呈球形,透明无色,大多分布在0~10 m水层中,卵径为0.95~1.25 mm,生殖季前期较大,之后逐渐变小。油球一个,球径为0.25~0.30 mm。颜色为极淡的米黄色,卵黄间隙小,卵子发育快,水温不同时需要的孵化时间也不同。
不同地区、不同种群鲐鱼的产卵期会有较大差异。台湾海峡北部外海的鲐鱼产卵期为1—5月,近海为3—5月;闽南、粤东近海地方群系的产卵期较长,几乎全年都进行产卵,4—5月为产卵盛期[10];对马暖流群系鲐鱼的产卵盛期为3—5月[11];太平洋群系鲐鱼的产卵期为1—6月[12],产卵高峰期为3—4月。
1.2 年龄生长及性成熟
鲐鱼性成熟叉长具有性别和地理区域的差异。黄海、东海鲐鱼性成熟年龄一般为2龄,少数个体1周岁即性成熟;日本海鲐鱼2龄时已有相当多的个体达到性成熟,然而大量鲐鱼性成熟的年龄则为3龄[13]。黄海鲐鱼初次性成熟叉长一般为250 mm[14];台湾海峡鲐鱼性成熟的最小叉长雌性为210~220 mm,雄性为200~210 mm[10]。闽南—粤东渔场的鲐鱼,性成熟的最小叉长和年龄均比闽中—闽东渔场的小,说明闽南—粤东地方群系的鲐鱼性成熟早于东海群系。
1.3 摄食
中国沿海鲐鱼摄食种类广泛,饵料生物包括24个类群50多种[15]。鱼类对饵料具有选择性,同一种鱼类的饵料在不同海域中有所不同,所以在不同海域鲐鱼的食物自然就不同。黄海区叉长35 mm以上的当年生鲐鱼幼鱼的饵料生物以鱼类、头足类和甲壳类为主,成鱼摄食对象以太平洋磷虾和鳀为主;福建近海的鲐鱼对饵料生物无明显的选择性,以浮游甲壳类和稚幼鱼为主;而南海北部鲐鱼的胃含物则以浮游动物和海底沉积物中的有孔虫类和翼足类为主[15]。研究表明,鲐鱼生物学特性不同的原因与种群、温度、盐度及各海域的资源开发利用情况等外界环境因素相关。如北太平洋公海海域鲐鱼最大叉长远大于东海南部和西非近海,最小叉长又远小于闽中、闽东渔场,且叉长优势组偏小,此外,8—10月渔获个体呈现逐月显著增大的趋势,这表明此时该海域饵料丰富,鱼体增长迅速[16]。
1.4 种群划分
目前,针对各海域鲐鱼种群的划分存在争议,不同学者的划分依据和标准各不相同。国内多数学者将中国海域的鲐鱼划分为3个种群:东海西部种群(黄海至闽中渔场)、五岛西部种群和闽南—粤东种群[15]。有日本学者将分布于东海不同越冬场的鲐鱼统称为对马暖流群系,但也有日本学者将在东海南部产卵,夏季到达东海北部及黄海,冬季洄游至东海南部的鲐鱼划归为东海西部群系,将分布于东海中部以北至山阴海域的鲐鱼划归为九州西部群系[17]。根据鲐鱼的生物学特性,有学者认为,日本海鲐鱼(包含在九州西岸等邻近海域的发生群)都作为对马暖流的一个群系较为妥当[18]。张丽艳等[19]利用AFLP分子标记的方法,对闽东和闽南渔场的鲐鱼种群进行了划分;Zeng等[20]利用微卫星技术,对东海和南海的鲐鱼种群进行了划分。综上所述,目前研究者对鲐鱼种群的划分仍存在争议,将来可采用鱼类框架形态判别、分子遗传判别等技术,扩大产卵场样本的覆盖范围,进行种群的准确有效划分。而本作者则认为,中国近海的鲐鱼有东海种群、闽南—粤东沿海地方种群、珠江口外海种群、清澜外海种群等4个种群[21]。
2 鲐鱼的早期生活史与环境的关系
2.1 鲐鱼的早期生活史研究
鱼类早期生活史研究是鱼类自然资源繁殖保护和养殖业苗种培育的基础[22],主要涉及卵和仔鱼发育、仔鱼最佳饲养条件、饵料密度、营养、生长、临界期、饥饿、捕食、环境耐力和毒性反应等。以往对鲐鱼的研究中多关注环境与成鱼资源间的关系,但每年的补充量主要由早期的存活决定,海洋环境因素的细微变化将对鱼类生命周期中最脆弱的鱼卵及仔幼鱼的生长、成活直至种群的补充都产生影响[23-24],因此,对于鲐鱼早期的生活史研究至关重要。
在鲐鱼早期生活史研究方面,国外学者起步较早,且大多在实验室进行观察研究,Hunter等[25]通过实验室饲养的方法对太平洋鲐鱼的早期生活史进行了研究,得到了鱼卵孵化时间、幼鱼变态时间与水温的相关关系,研究发现,其生长速度较快、新陈代谢较高(图4)。Yamada等[26]在实验室进行了鲐鱼繁殖力的试验,得到了产卵时间、分批产卵次数、总产卵量的数据,研究发现,产卵亲体的营养情况影响产卵的繁殖力。然而,实验室进行的饲养试验费时费力,研究资源和方式等均受限制,相对来说建立模型进行模拟的可操作性和经济性较为适宜。Yukami等[27]根据渔获量统计和生物特征数据资料,估算出了东中国海鲐鱼的产卵场,预计2—4月的鲐鱼产卵场在东海中部、南部及九州岛的西部海域,5月的产卵场在东海中部,以及对马岛和九州岛以西海域,6月的产卵地移至北部地区的对马岛和日本海西部海域。

实线为目前的数据,点为5个试管中卵孵化50%的温度,虚线为Watanabe(1970)的数据。由Zweifel和Lasker(1976)提出的一般方程,拟合孵化50%的值。
因仔鱼期阶段的生长率对翌年的资源补充量有着极大的影响,Kamimura等[28]根据采集到的鲐鱼仔稚鱼,通过分析耳石年龄,研究其生长规律,发现海表温度与幼体日平均生长率间有较明显的相关关系。较高的水温导致太平洋鲐鱼在4月份生长速度加快,幼鱼期缩短,这表明4月份鲐鱼生长率是其生活史上一个主要的指标。
2015年3月—2016年7月通过对日本大岛太平洋沿岸的鲐鱼资源进行调查,Taga等[29]采用广义线性混合模型(generalized linear mixed model,GLMM)研究了温度和饵料密度对幼鱼近期生长率的影响,结果表明,幼体对饵料需求的增加可能导致其生长依赖于饵料密度。
在鲐鱼早期的生活史过程中,Bartsch等[30-32]将个体模型(individual based model,IBM)应用到大西洋东部鲐鱼的研究中,并开发了物理-生物输运模型,以预测其早期生活史阶段的存活率。李曰嵩等[1,33-34]采用FVCOM(finite volume coastal ocean model)物理模型,利用基于个体模型参数化东海鲐鱼早期的生物过程,构建了物理-生物耦合的东海鲐鱼生长初期的生态动力学模型,以模拟研究鲐鱼鱼卵和仔鱼的运输和分布等情况,结果发现,物理环境对鲐鱼早期生活史影响较大,4月份的鲐鱼仔幼鱼存活率决定了鲐鱼资源补充量,并为深入研究气候变化对鲐鱼资源后续补充量的影响提供了数据参考。
2.2 外界环境变化对鲐鱼的影响
2.2.1 水温 鱼类对海洋环境的适应性和局限性决定了鱼类的洄游、分布移动,鱼类资源的分布及其时空变动与海洋环境紧密相连[35]。海洋环境变化势必对鱼类资源变动、时空分布产生重要的影响,受海洋环境变化的影响,东海鲐鱼资源的时空分布就具有非均匀性和动态性的特点,其中,水温作为影响鱼类的物理因素之一,会对鲐鱼产卵、生长、洄游和补充量产生影响。苏杭等[36]利用中国大型灯光围网的鲐鱼生产统计数据(1999—2007年)并结合海洋遥感数据,分析了渔汛期间鲐鱼栖息地的适宜海洋表面温度(sea surface temperature,SST)范围,并探讨了SST改变后鲐鱼栖息地的变动情况。结果表明,SST的升高可能对近海鲐鱼栖息地产生较大影响。李曰嵩等[34,37]基于个体模型对东海鲐鱼补充量的年际变动进行模拟,结果发现,4月份的水温是影响鲐鱼资源补充量的一个重要原因,该月水温升高会增加鲐鱼资源补充量。官文江等[38]研究发现,东海鲐鱼在越冬、产卵洄游过程中,鲐鱼资源量与海表水温具有相关关系,认为海表水温的变化会影响鲐鱼的洄游路线,并引起鲐鱼资源的不均匀分布。1997年,Nishida等[39]对1969—1978年日本海鲐鱼种群产卵与补充量的关系进行了分析,发现在此期间出现的厄尔尼诺事件使得海水水温升高,并提出东海海表水温上升有利于提高鲐鱼的资源补充量。Hiyama等[40]通过分析日本大中型围网渔业的渔获量数据,研究了东海和日本海的鲐鱼资源状况,讨论了海水温度与鲐鱼产卵量波动的关系,认为产卵成功率与海洋表面温度呈负相关关系(图5,图中黑点表示1985—1999年的年份)。

2—6月的平均值(30°N、125°E)。
2.2.2 盐度和海表面高度 除海水温度外,基于盐度、海表面高度两个环境因子的权重会对东海鲐鱼栖息地产生较大影响,易炜等[41]研究表明,随时间的推移,即所在的月份不同,海水温度、盐度、海表面高度等环境因子会导致鲐鱼渔场分布差异。官文江等[42]研究了海洋环境对东、黄海鲐鱼灯光围网捕捞造成的影响,发现渔场中的海表水温、叶绿素、平均海面高度距平等多个环境因子与CPUE存在联系,月光的亮度、风速等也会对鲐鱼的捕捞产生一定的影响。
2.2.3 气候条件 同样,李曰嵩等[43-44]基于个体模型,探讨了Alice台风对东海鲐鱼鱼卵和仔鱼的输运、存活率和补充量的作用,结果发现,由于台风自身移动速度快、时间短的特点及气旋间的彼此抵消等因素,Alice台风对鲐鱼仔鱼的输运分布造成的影响较小,但台风可使鱼卵和仔鱼向深水处移动,最终会使仔鱼存活率下降。杨胜龙等[45-46]基于GAM模型对阿拉伯海鲐鱼渔场时空分布与海洋环境的关系进行了研究,发现空间因子和环境因子会对渔场产生较大的作用,按照对鲐鱼渔场影响程度的重要性,依次为海面高度、经纬度位置、海表温度和叶绿素浓度。
厄尔尼诺、拉尼娜事件对气候、海洋环境产生的影响错综复杂,故研究其对渔业资源的影响则更难,且不同的学者曾有不同的结论。郭爱等[47]研究了厄尔尼诺、拉尼娜事件对鲐鱼栖息地的影响,结果表明,中国近海鲐鱼栖息地适宜性与厄尔尼诺、拉尼娜异常气候事件的发生有密切关系,且随其发生的强度不同而产生变化,当发生超强厄尔尼诺事件时,鲐鱼的适宜栖息地面积减小,鲐鱼CPUE骤降,强拉尼娜事件则反之。官文江等[38]则提出,拉尼娜事件会对东海不同海域鲐鱼资源产生不利影响,但原因还有待进一步研究证实。
3 应用于鲐鱼生活史及资源预测的相关模型
在鲐鱼的早期生活过程中,鱼卵和仔幼鱼基本处于被动漂流状态,利用模型可以较好地对鲐鱼的早期生活史过程进行模拟研究。鱼类巨大的资源量、广泛的空间分布、难以准确采样等特点,加之不断变化的全球气候和人类对海洋系统的日益破坏,有可能从根本上改变海洋系统的动态,这些都使生态学家较难进行现场种群动力学的研究,为此生态动力学模型在鱼类资源研究中扮演了重要的角色[48]。动力学模型将物理和生物模型耦合,可以从产卵、生长、死亡、补充量、输运与丰度分布、环境变化等对鲐鱼的鱼卵和仔幼鱼影响进行模拟研究。动力学模型需要应用物理模型来驱动生物模型,本研究中主要综述了采用物理模型及生物、生态模型对鲐鱼生活史及资源状况的研究进展,可为今后在模型的选择方面提供帮助。
3.1 物理模型
海洋环流的变化会改变仔幼鱼和营养物的运输方向,水温的变化会影响鱼类的繁殖和生长速度,因此,驱动生态动力学模型的海洋模型选择至关重要。海洋模型按水平网格的离散方式及所使用垂向坐标系的不同分为几个发展阶段,最早出现且还在使用的海洋模型是基于原始方程的低阶精度的有限差分模型,它在水深方向用z坐标系表示,例如HOPS(harvard ocean prediction system)、MOM(GFDL modular ocean model)、POP(parallel ocean model)和NCOM(NCAR community ocean model)等模型在某种程度上都可以认为是有限差分模型的改进版。当Sigma坐标系用于海洋模型的水深方向后,出现了POM(princeton ocean model)、ECOM(estuarine coastal and ocean model)和ROMS(regional ocean model)等模型。海洋模型中开始运用非结构化网格技术后,出现了SEOM(spectral finite element ocean model)和FVCOM(finite volume coastal ocean model)模型[49]。21世纪以来,把目前最先进的物理知识、数值方法和数据同化技术集为一体的TOMS(terrain—following ocean modeling system)模型开始发展。FVCOM、HYCOM(HYbrid coordinate ocean model)和ROMS模型是几种目前常用于渔业生态动力学的海洋模型。
3.1.1 FVCOM模型 该模型是无结构网格、有限体积、三维原始方程的海洋模型。模型包含动量方程、温盐守恒方程及状态方程,FVCOM模型是能够并行计算、模块化且适用于不同需求的模型。其优势是运用了结合有限元法的网格几何灵活性和有限差分法的简单离散计算的有限体积法。Chen等[48]利用高分辨率、非结构网格、有限体积沿岸海洋模式(FVCOM),建立了东中国海的海洋模型(the unstructured grid finite-volume coastal ocean model for East China Sea,FVCOM-ECS),其分辨率为0.1~15.0 km,垂向分为40层,可为东海鲐鱼IBM模型提供高分辨率的三维流场、温盐场、湍流扩散参数[1]。Huret等[50]利用FVCOM模型初步建立了大西洋鳕生长发育与环境变异间的动力学模型,李曰嵩[1]也利用FVCOM模型构建了东海鲐鱼早期生活史生态动力学模型。
3.1.2 HYCOM模型 该模型是一种采用混合坐标系的三维水动力模型,模型克服了传统模型系统设计及垂直离散化的局限性。其设计初衷就是为了应用的区域范围更加广泛,且在模拟、预报等方面的表现力更加突出。HYCOM模型经过多年发展,已逐渐发展为比较完备的模型系统,在珊瑚Coralliumsulcatum[51]、冰冷珊瑚Lopheliapertusa[52]幼体等许多研究中得到了较好的应用。
3.1.3 ROMS模型 该模型是一类三维、自由表面、地形跟随的海洋数值模型,被广泛应用于海洋及河口地区的水动力、水环流模拟和海洋环境预报中。在海洋生物,如南极磷虾Euphausiasuperba(仔稚期)[53]、侧纹南极鱼Pleuragrammaantarctica[54]、冲浪蛤Spisulasolidissima[55]和欧洲沙丁鱼Sardinapilchardus[56]等研究中都进行了应用。
3.2 基于个体的生物和生态模型
许多鱼类种群模型依赖于统计估计的参数,如生存率、死亡率和发育时间,这些参数会因环境改变而以更为复杂的方式发生变化。基于个体生物模型(IBM)可以解释环境影响个体的生命活动,以及个体与环境间的相互作用,该模型包含了关于自下而上底层环境影响鱼类生态过程,这种方法也被认为是研究鱼类生态过程的唯一合理手段。
基于个体模型是以众多的生物个体为模拟对象,并考虑个体间差异及环境条件的时空变化对个体发育的影响,在此基础上统计出种群水平的属性[57]。所以对于不同的研究对象,环境因子和参量不相同。基于个体模型可以模拟物种的生物学特点和海洋物理情况,此模型把种群当作个体的集合体,对鱼类种群的生长、输运、时空分布、洄游、滞留、产卵、群体生物量等进行研究。基于个体模型侧重于生物、生态耦合水动力,模拟研究物理环境,例如海流、海温、盐度、光照等变化对海洋生物生存、死亡、运输和分布的作用。目前,个体模型主要应用在一些商业价值高的鱼类,如大西洋鳕Gadusmorhua、太平洋鲱Sprattussprattus等,研究内容包括探讨鱼类生活史及其与环境间的关系,以及寻找最适栖息地[58]等方面。
Bartsch等[30-32]构建了基于环流HAMSO模型提供的物理场,对太平洋鲐鱼早期生活史阶段进行模拟生长和输运的IBM模型,并于2001年利用该模型对仔幼鱼输运分布进行模拟,结果发现,较温暖水域对浮游时期的卵、幼鱼和幼鱼后期的存活有利。2004年该研究团队用此模型模拟鲐鱼鱼卵、幼鱼和幼鱼后期的运输、生长和死亡率,预测北大西洋东部鲐鱼早期生活史阶段的年存活率,同时,还研究了改变物理场对鲐鱼存活率产生的影响,并建立北大西洋东部鲐鱼仔鱼的生态动力学模型。
李曰嵩等[1]根据鲐鱼生长初期的生活环境和生长特性,利用FVCOM模型构建了基于个体的东海鲐鱼早期生活史物理-生物耦合生态动力学模型。利用该模型验证在台湾东北部产卵场的鱼卵、仔幼鱼是向对马海峡方向进行输运,并推算从东海南部产卵场到育肥场的仔幼鱼数量比例,研究了产卵场对各育肥场的补充性和联通性,分析了对输运分布和存活率产生影响的动力学因素;此外,该团队还研究了在异常气候及鲐鱼产卵场发生变化时,对鱼卵、仔幼鱼的输运分布及资源补充量产生的影响,分析了其动力学规律,并利用该模型对仔幼鱼集群现象及成鱼渔场形成动力学原因进行了分析。2014年,该学者模拟了鲐鱼幼鱼在东中国海的扩散,并通过一系列敏感性试验,探究了东海鲐鱼幼鱼存活率、扩散路径受台风、水温、孵化场地的影响[59]。这些研究均表明,区域环流、垂直分层和季节变化会直接影响幼鱼的分布位置和数量[60]。
李向心[57]基于IBM模型模拟了鳀鱼种群对环境变化和捕捞活动的响应,研究了温度对黄渤海鳀鱼种群的分布和生物量等的影响;王玉衡[61]基于IBM模型研究了黄海物理环境对鳀鱼种群数量的动态影响,探讨了黄海较复杂的水文环境对鳀鱼生活史各个阶段的影响,也为未来其他物种的IBM模型开发提供了科学参考。
3.3 基于群体的生物和生态模型
伴随着科学技术的迅速发展,国内外涌现出了许多基于个体或群体的生物、生态模型。目前,已有多个模型被运用到鲐鱼生态过程模拟的研究中,常见的几种生物、生态模型列于表1,并与IBM模型进行了比较。
根据2006—2015年中国鲐鱼生产统计的鲐鱼产卵场和索饵场的相关数据,易炜等[62]基于剩余产量模型进行分析,建立了基于环境因子的东、黄海鲐鱼生物经济模型,研究了环境因子对鲐鱼资源造成的影响。薛艳会等[63]构建了基于灰色系统的对马群系鲐鱼资源丰度预测模型,并对鲐对马群系资源丰度进行预测,利用1973—2016年日本西海水产研究所提供的鲐鱼对马群系的资源量与渔获量数据,结合产卵场、索饵场的海表面温度等数据分析验证,得到了对马群系资源丰度最优预测模型。王易帆等[64]进行了近海鲐鱼资源丰度的3种中长期预测模型的比较,并利用多元线性回归模型、BP神经网络模型和灰色系统GM(1,N)3种模型建立了资源丰度指数和环境因子间的资源丰度预测模型,找出了最佳模型。高峰等[65-66]在对提升回归树模型建立、算法求解和模型参数的选择进行解释说明的基础上,构建了对东、黄海鲐鱼渔场预报情况模拟研究的提升回归树模型,为东、黄海鲐鱼的渔场预报和渔情分析提供了科学依据。宋利明等[67]采用分位数回归的方法,建立了毛里塔尼亚海域鲐鱼CPUE与叶绿素a浓度、海表面温度、海表面盐度的关系模型,利用广义加性模型(GAM)来评价环境因子的影响程度。
有学者基于贝叶斯原理构建了鲐鱼资源模型。徐香香等[68]构建了西北太平洋鲐鱼渔场预报模型,综合预测鲐鱼的渔场数据;王从军等[69]构建了东、黄海鲐鱼贝叶斯生物经济模型,对鲐鱼资源量和渔业短、中、长期利润的变化进行模拟;李纲等[70]构建了基于贝叶斯方法的剩余产量模型,研究了东、黄海鲐鱼当前的资源开发利用状态,模拟估算了不同收获率水平下未来5年鲐鱼渔业资源量。
从表1可见,每种模型都有其优缺点,应用单一模型不能克服其自身的不足。建立多种模型进行比较分析,能够为鲐鱼资源量的更精确预测提供科学依据。
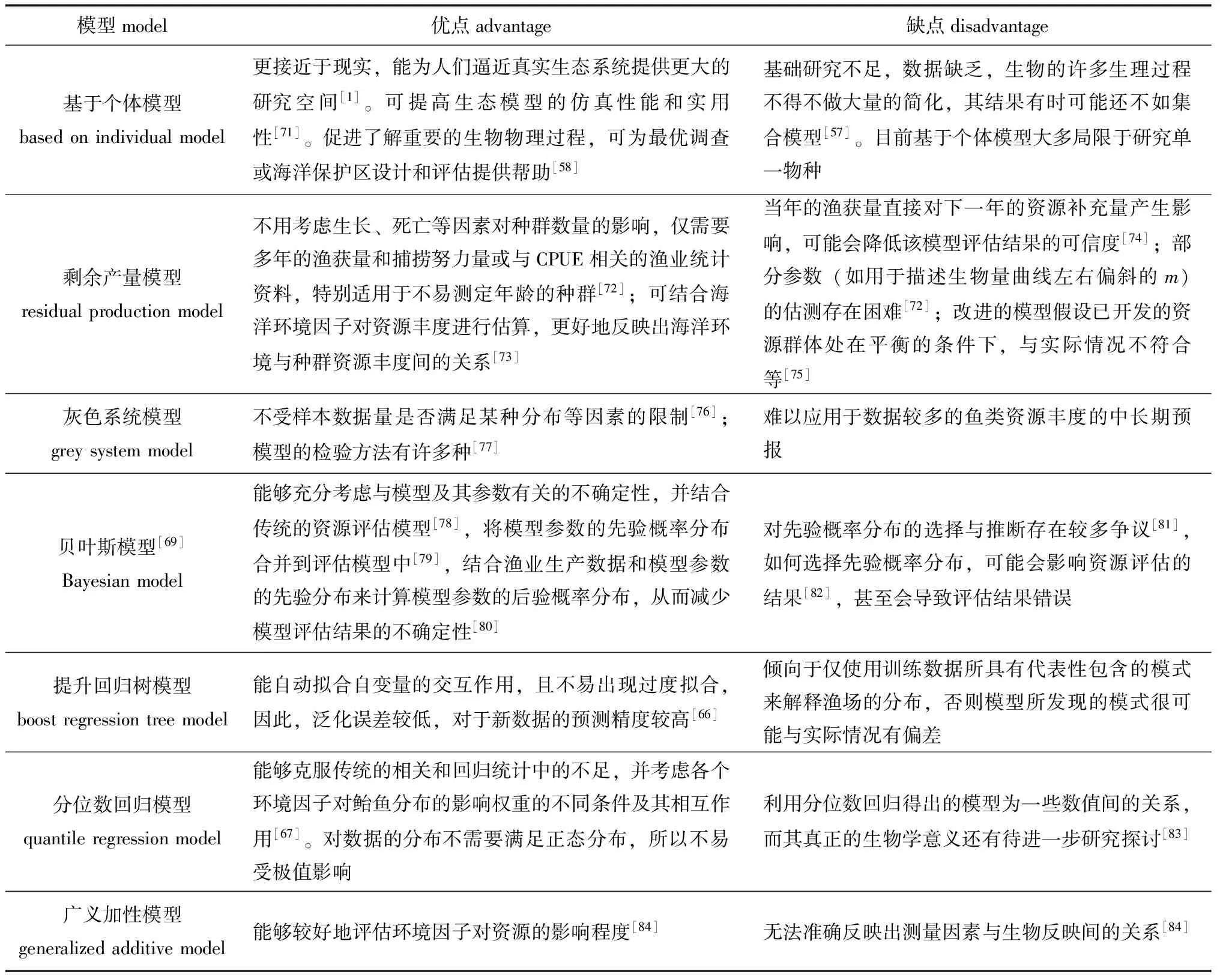
表1 鲐鱼生态过程模拟研究中常见的几种生物和生态模型
4 存在问题及展望
4.1 鲐鱼生活史及模型应用中存在的问题
鲐鱼分布广泛且经济价值高,但外界环境变化对其种群资源补充量和时空分布的影响较大。尽管有学者进行了环境对鲐鱼资源影响的相关研究,但仍存在以下一些问题:
1)鲐鱼的生物学研究方面。目前,虽然已有大量关于鲐鱼的研究,但大部分集中在鲐鱼的生物学特性、渔场分布、资源量评估与管理等方面,而对于鲐鱼早期生活史的研究相对较少,鲐鱼早期的试验数据则更少。这就导致在鲐鱼生态动力学模型研究中,缺乏鲐鱼早期生活史阶段的参数,使构建的模型降低了可信性。今后应当加大鱼类基础生物学研究的力度。
2)鲐鱼的种群划分方面。鲐鱼的分布、洄游范围广泛,不同群体间存在基因上的交流,国内外学者分别有不同的群体划分依据,目前还未达成共识,且一直存在争议。今后可以利用鱼类框架形态和分子遗传等判别技术,增加产卵场样本采集覆盖面以探究其种群,加强种群的科学、精确、统一标准划分,以便更有针对性地对鲐鱼种群进行研究。
3)环境对鲐鱼资源的影响方面。鲐鱼资源量受气候和海洋环境的影响显著,海洋物理环境对鲐鱼的洄游、分布、繁殖与生长有重要影响,这方面的研究大多来源于调查和捕捞数据,利用数理统计的方法进行资源量与环境因子相关性的研究,较难揭示资源变动的真实原因。气候变化导致的海洋环境变动同样会对鲐鱼资源量产生影响,通过研究厄尔尼诺和拉尼娜这种重大气候变化对鲐鱼资源量的影响,不同学者得出的结论存在差异,有的甚至截然相反。未来要加强鲐鱼生态动力学的基础研究,以揭示气候和物理环境变动对鲐鱼补充量影响的动力学本质。
4)鲐鱼的生物和生态模型应用方面。鱼类生态动力学模型是在海洋模型与鱼类生物模型基础上交叉发展起来的,其核心问题是海洋物理过程与鱼类生物过程的相互作用和耦合,在国外应用十分普遍。在国内,由于专业背景所限,鱼类学家对物理海洋相关知识了解不够深入,对海洋模型的模拟就更难掌握,导致基于个体模型的研究应用在国内不多。另外,虽然鲐鱼模型应用方面已经取得了一些成果,但由于鲐鱼生命活动较为复杂,在模型中需要考虑多种要素,模型的准确性需要提升。今后需加强鱼类学和海洋物理学的交流合作,将最新的研究成果应用到模型中来,利用模型加强对鲐鱼早期生活史的研究,促进中国鱼类生态动力学的发展。另外,在鲐鱼的早期生活过程中,鱼卵和仔幼鱼基本处于被动漂流状态,进行模型模拟研究更加可控。未来研究可以利用模型将鲐鱼早期生活史阶段参数化以研究资源补充过程。
4.2 未来重点研究方向
科学养护和管理近海重要渔业资源,是实现中国海洋渔业可持续发展的重要举措,也是实现海洋生态文明的重要抓手。目前,鲐鱼已成为中国近海的主要经济鱼种,是灯光围网渔业重要的捕捞对象。因此,科学养护和管理鲐鱼资源,确保其资源可持续利用,成为包括中国在内的周边国家和地区共同关注的国际渔业问题。未来应在以下几个方面重点开展研究:
1)鲐鱼对全球气候变化生态响应研究。鲐鱼资源量易受全球海洋环境等因素的影响,年间资源量波动较大,相关研究可利用模型评估全球气候变暖及海洋环境变化对鲐鱼资源量的影响,掌握全球气候和环境变化对鲐鱼资源的影响机制。
2)鲐鱼中长期预测及鲐鱼渔业发展战略。结合全球气候和环境变化趋势对鲐鱼资源影响的机制进行研究,预测短期(各年度)和中长期(3~5年)中国近海鲐鱼的资源量,并对近海鲐鱼渔船发展规模、生产安排和规划,以及合理布局中国近海鲐鱼渔业,提出相应的建议。
3)近海鲐鱼的对外谈判战略。鲐鱼是中日韩三国共享资源,目前在周边渔业合作和谈判中是主要渔业种类。今后应在鲐鱼资源中长期预测的基础上,探讨其资源量的空间分布,分析其捕捞方案及中国在对外渔业谈判中的策略,积极争取更多的海洋渔业权益。
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
中学历史教学(2016年4期)2016-11-11
中国社会历史评论(2016年2期)2016-06-27
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
应用海洋学学报(2014年1期)2014-11-22
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
中国现当代社会文化学术沙龙辑录(2013年0期)2013-10-24
中国烟草学报(2012年2期)2012-04-09
