
铜胁迫对丝瓜幼苗色素含量、光合特性及渗透调节系统的影响
2021-09-13 02:36蒋英郭红伟
天津农业科学 2021年7期
关键词:丝瓜
蒋英 郭红伟
摘 要:為探讨丝瓜幼苗对铜的耐受范围,以‘美秀丝瓜品种为试材,借助营养液沙培的方式,研究不同浓度的铜胁迫(Cu2+ 0,5,15,30,50 μmol·L-1)对丝瓜幼苗光合色素、光合参数及渗透调节物质含量的影响。结果表明:随着铜浓度的增加,丝瓜幼苗叶片叶绿素a、叶绿素、类胡萝卜素含量呈先升后降的趋势,均于铜浓度5 μmol·L-1时达到最大值;净光合速率、气孔导度和胞间CO2浓度均呈现逐渐降低的变化趋势,其中铜浓度5 μmol·L-1处理的净光合速率、蒸腾速率和胞间CO2浓度均与对照差异不显著,但其他处理均显著低于对照,蒸腾速率呈先升后降的趋势且于铜浓度5 μmol·L-1达到最大值;可溶性糖含量呈现先升高再降低的单峰变化规律,且在铜浓度5 μmol·L-1时值最大;游离脯氨酸含量呈现显著上升的趋势。综上,铜浓度不高于5 μmol·L-1时,能提高丝瓜幼苗光合色素含量,保持较高的净光合速率,同时通过渗透调节物质的调节保护叶片细胞免受重金属铜毒害。
关键词:丝瓜;铜胁迫;光合色素;光合参数;渗透调节
中图分类号:S642.4 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2021.07.002
Abstract: To explore the tolerance range of Luffa cylindrical seedlings to copper,the effects of different concentrations of copper stress (Cu2+ 0,5,15,30,50 μmol·L-1) on photosynthetic pigment, photosynthetic parameters and osmotic adjustment substance content of Luffa cylindrica seedlings were studied by using 'Meixiu' varieties as test materials in the method of nutrient solution sand culture. The results showed that, the contents of chlorophyll a, chlorophyll and carotenoid were increase first and then decrease following with the increased copper concentration, which the maximum value were all appeared at the copper concentration of 5 μmol·L-1. With the increase of copper concentration, the net photosynthetic rate, stomatal conductance and intercellular CO2 concentration of Luffa cylindrica seedlings decreased gradually,in which the treatment of 5 μmol·L-1 copper concentration had no significant difference with the control but the other treatments were significant lower than the control; the transpiration rate was rising first and then decreasing, and the maximum value was appeared at the copper concentration of 5 μmol·L-1; following with the copper concentration, the soluble sugar content of Luffa cylindrica seedlings showed a single peak change and the maximum value was obtained at the copper concentration of 5 μmol·L-1, while the free proline content was significantly increased. In summary, when the copper concentration was not higher than 5 μmol·L-1, the photosynthetic pigment content of Luffa cylindrica could be increased,the net photosynthetic rate were maintained at the level consistent with the control, and the leaf cells were protected from the heavy metal copper toxicity by the regulation of osmotic regulators.
Key words: Luffa cylindrica; copper stress; photosynthetic pigment; photosynthetic parameters; osmotic regulation
铜是高等植物生长发育必需微量元素之一,是超氧化物歧化酶、多巴胺-β-加氧酶、细胞色素氧化酶等多种酶的重要组分[1],同时铜又参与作物产量、品质、形态建成等众多生理代谢过程,是植物生长发育过程中必不可少的营养元素。但环境中铜含量过高时会导致植物体遭受铜毒害,如根系生长不良、根系变短、根条数减少、株高下降、叶片失绿、分蘖减少、种子发芽率降低、PSⅡ反应中心的电子浓度下降[2],细胞器功能受损[3],产量降低、品质下降、品种退化[4]。当前有关铜胁迫研究主要集中小麦、玉米、水稻等粮食作物[5],有关蔬菜作物方面的铜污染研究较少,有关丝瓜铜污染方面的研究尚未见报道。丝瓜果实是人们较为喜食的重要蔬菜之一,借助日光温室可以实现周年生产供应。但是在丝瓜的周年生产过程中,除草剂、农药、化肥的大量使用,使用被工业废水污染的水源进行灌溉等因素均会使土壤铜含量升高。铜被植物吸收后,在植物体内具有累积性,当达到一定阈值后就会影响植株的正常生理代谢,甚至对人类的健康造成危害。因此,本试验借助营养液沙培方式,研究不同浓度铜胁迫对丝瓜幼苗光合色素、光合参数及渗透调节系统的影响,以期为蔬菜生产、铜污染防治提供理论依据,为铜毒害的作用机理提供数据参考。
1 材料和方法
1.1 供试材料
供试丝瓜品种为‘玉秀,由京研益农种业科技有限公司提供。
1.2 试验设计
于2020年2月25日播种育苗,4月3日选取长势基本一致的3叶1心丝瓜幼苗,栽培于长3 m、宽30 cm、高15 cm的栽培槽内,每个栽培槽定植20株,栽培槽内填充洗净且消毒过的河沙,栽培槽底部有排水阀。试验共设置1个对照和4个处理,以Hoagland营养液为对照,在Hoagland营养液基础上添加CuSO4·5H2O,使营养液中Cu2+浓度分别达到5,15,30,50 μmol·L-1,作为4个处理。处理和对照均设置3次重复,1个栽培槽作1次重复,共15个栽培槽。栽培槽内浇灌相应的营养液后,胁迫处理7 d后对相关指标进行测定。
1.3 测定项目及方法
于2020年4月10日上午9:00—10:00测定丝瓜叶片净光合速率,随后取样测定叶绿素含量。叶绿素含量、净光合速率、游离脯氨酸和可溶性糖含量等指标的测定均选用丝瓜幼苗第2片真叶。其中,采用乙醇丙酮浸提法测定叶绿素含量[6];采用美国LI-COR公司生产的LI-6400光合仪测定叶净光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)、胞间CO2浓度(Ci);采用蒽酮比色法测定可溶性糖含量[7],参照茚三酮-磺基水杨酸法测定脯氨酸(Pro)含量[8]。
1.4 数据处理
试验数据使用Microsoft Excel 2007进行处理和作图,用SPSS13.0软件进行统计分析。
2 结果与分析
2.1 不同铜处理对丝瓜幼苗叶片光合色素含量影响
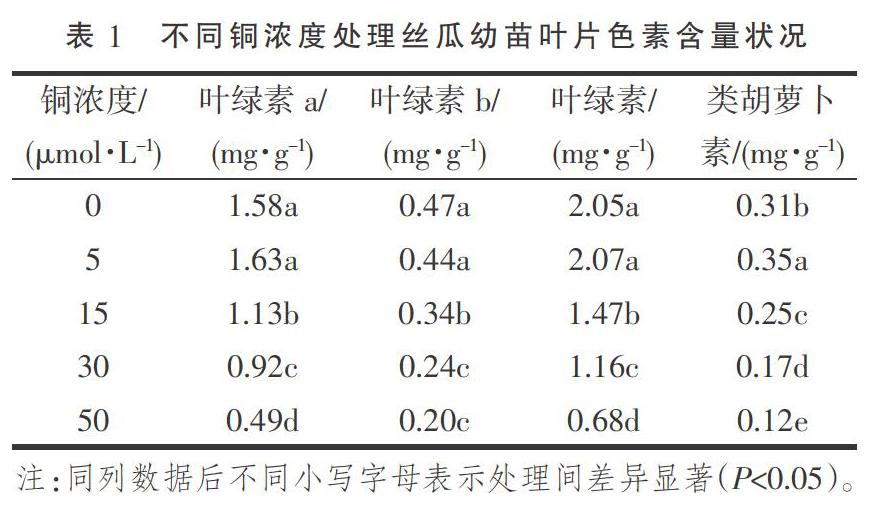
由表1可以看出,随着铜浓度的增加,丝瓜幼苗叶片叶绿素a和叶绿素含量表现出先升高再降低的单峰变化规律,最大值均出现铜浓度5 μmol·L-1时,与对照差异均不显著但均显著高于其他铜处理;叶绿素b呈下降趋势,其中5 μmol·L-1铜浓度处理与对照差异不显著,其他处理均显著低于对照;类胡萝卜素含量亦呈先升后降的单峰变化规律,且在铜浓度5 μmol·L-1时类胡萝卜素含量最大,较对照显著增加12.90%,而其他铜浓度处理均较对照显著降低。可见,低浓度≤5 μmol·L-1的铜处理能够促进丝瓜幼苗叶片光合色素的合成与提高,高浓度的铜处理则对其有抑制作用。
2.2 不同铜处理对丝瓜幼苗叶片光合参数的影响
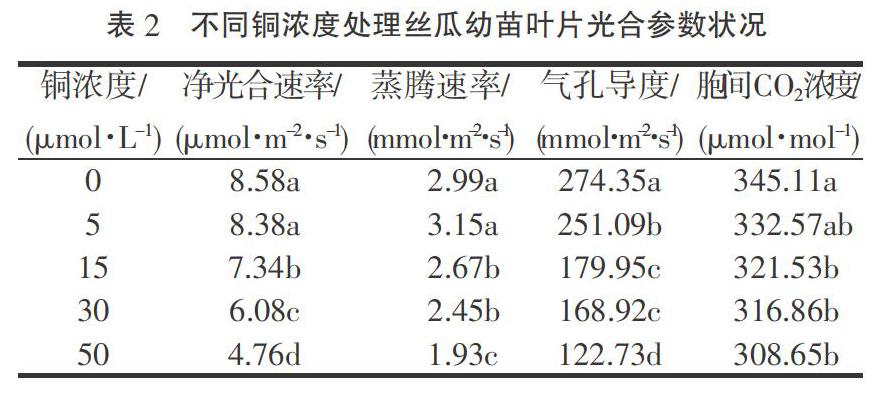
由表2可知,随着铜浓度的增加,丝瓜幼苗净光合速率逐渐降低,但在铜浓度5 μmol·L-1时与对照差异不显著,而铜浓度15,30,50 μmol·L-1时较对照分别显著降低14.45%,29.14%,44.52%;丝瓜幼苗叶片气孔导度和胞间CO2浓度的变化规律与净光合速率一致,亦呈下降趋势,其中各铜处理气孔导度均显著低于对照,而胞间CO2浓度在铜浓度5 μmol·L-1时与对照差异不显著,其他铜处理均显著低于对照;丝瓜幼苗叶片蒸腾速率呈先升高后降低的趋势,其中铜浓度5 μmol·L-1处理达到最大值,但与对照差异不显著,其他铜浓度处理均显著低于对照。
2.3 不同铜浓度处理丝瓜幼苗渗透调节物质状况
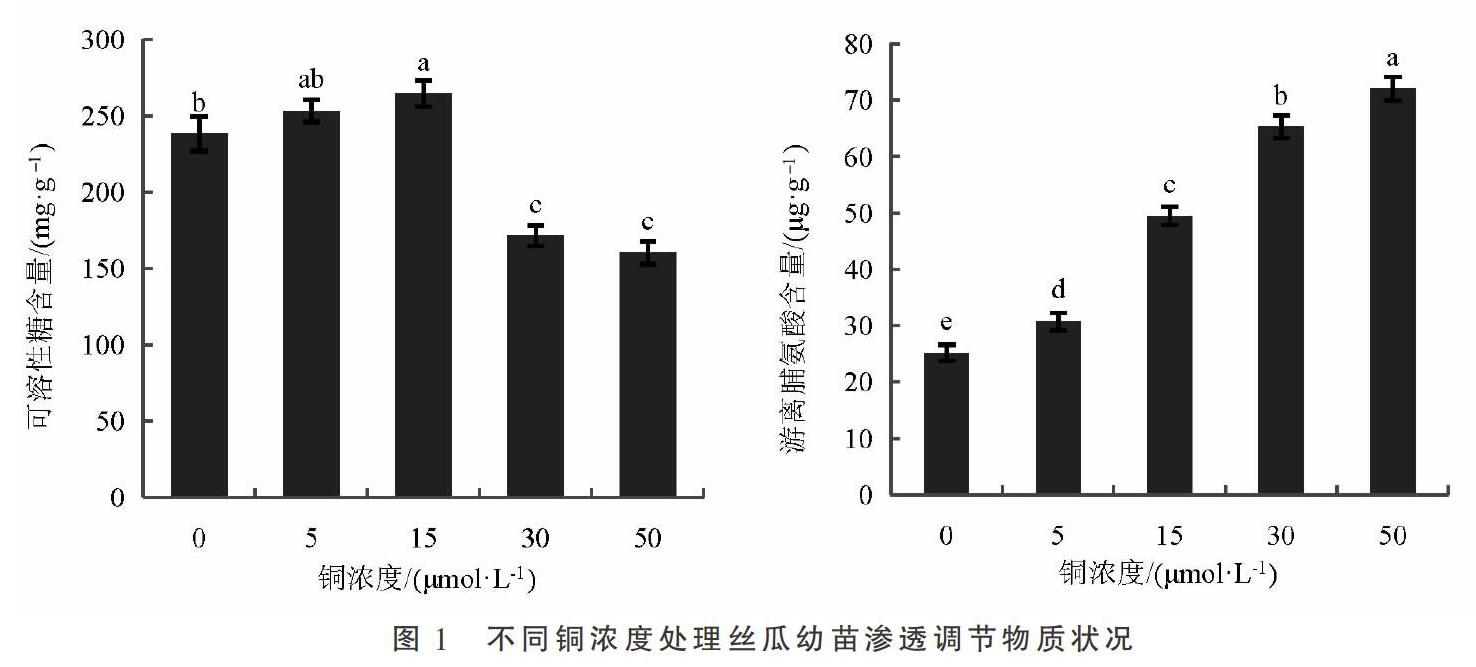
由图1可知,随着铜浓度的增加,丝瓜幼苗可溶性糖含量呈现先升高再降低的单峰变化规律,铜浓度15 μmol·L-1时达到最大值,较对照显著增加11.11%(P<0.05),铜浓度5 μmol·L-1时虽高于对照但与对照差异不显著,30,50 μmol·L-1铜浓度处理均显著低于对照,较对照降低28.08%,32.71%;丝瓜幼苗游离脯氨酸含量呈现显著上升的趋势,铜浓度50 μmol·L-1时较对照增加186.30%。
3 结论与讨论
叶绿素含量高低除受自身生理特性及遗传因素有关外,还受外界环境因子的制约,当植物受到逆境胁迫时,其含量会受到影响[9]。李永杰等[10]研究得出,Cu2+质量分数低于2.5×10-4时,可促进白蜡幼苗叶绿素含量的增加;Cu2+质量分数达到5×10-4及以上时,抑制叶绿素的生成。张自坤等[11]研究指出,随着铜浓度的增加,嫁接苗黃瓜和自根苗黄瓜叶绿素和类胡萝卜素含量均降低。本试验中,铜浓度低(≤5 μmol·L-1)时能够促进丝瓜幼苗叶绿素和类胡萝卜素含量的增加,这应该是因为铜作为叶绿素合成过程中某些酶的活化剂,有助于促进叶绿素合成酶活性的提高,故适量的铜有利于叶绿素的形成与稳定[12];铜浓度达到15 μmol·L-1及以上时,叶绿素与类胡萝卜素含量下降,即表现为铜胁迫的抑制作用,推测是因为铜浓度过高时Cu2+与叶绿体蛋白质上的巯基结合,使叶绿体失活而造成叶绿素含量下降[13]。
净光合速率是反应植株叶片光合能力强弱的重要生理指标[14]。本试验中,在铜浓度5 μmol·L-1时,丝瓜幼苗叶片净光合速率、气孔导度、胞间CO2浓度均低于对照,这表明此时净光合速率的降低是由铜胁迫引起气孔导度下降,CO2供应受阻,光合底物减少所致,也即气孔因素限制引起的净光合速率降低;当铜浓度15~50 μmol·L-1时,随铜浓度的增加,净光合速率、气孔导度继续降低,而胞间CO2浓度下降不显著,这表明此时引起的净光合速率的降低是非气孔因素所致,这可能是光合机构受损,光合磷酸化活性、核酮糖-1,5-二磷酸羧化酶(Rubisco)活性降低,叶绿体电子传递能力下降等因素引起的非气孔限制[15]。
可溶性糖和游离脯氨酸都是重要的渗透调节物质[16],对植物维持细胞完整性、抵御逆境胁迫有重要作用。本试验中,铜胁迫条件下,丝瓜幼苗叶片脯氨酸含量显著升高,且随铜浓度的增加而增大,这是因为脯氨酸能够协调原生质体内外的渗透平衡,调节膨压,以解除电解质的毒害[17],维持细胞的完整性;在铜浓度0~15 μmol·L-1时,可溶性糖含量随铜胁迫浓度的增加而增大,至铜浓度30 μmol·L-1及以上时,可溶性糖含量显著降低,这与徐磊[18]在小白菜铜胁迫上的试验结论一致。可溶性糖的变化规律可能是因为在低强度的铜胁迫下,淀粉等高分子碳水化合物合成减少而分解加快,形成包括蔗糖等在内的低分子量物质,但在高强度的铜胁迫下,细胞膜严重受损、电解质大量渗漏,阻碍可溶性糖含量的增加[19]。
综上,铜浓度不高于5 μmol·L-1时,能提高丝瓜幼苗光合色素含量,保持较高的净光合速率,同时通过渗透调节物质的调节保护叶片细胞免受重金属铜毒害。因此,在实际生产中,对植株补充微量元素铜时,其浓度不得高于5 μmol·L-1,对于铜浓度高于5 μmol·L-1的土壤也不适宜种植丝瓜,否则会影响丝瓜的正常生理代谢,最终影响丝瓜的产量和品质。
参考文献:
[1] 王镜岩. 生物化学[M]. 北京: 高等教育出版社, 2002.
[2] GUPTA A, SINGHAL C S. Inhibition of PS Ⅱactivity by copper and its effect on spectral properties on intact cells in Anacystis nidulans[J]. Environ.Exper.Bot, 1995, 35(4): 435-439.
[3] STRANGE J, MACBAUR M. Evidence for a role for the cell membrane in copper tolerance of Mimulus Guttatus Fischer ex DC[J]. New physiol, 1991, 119: 383-388.
[4] 张太平, 段昌群, 胡斌, 等. 玉米在重金属污染条件下的生态分化与品种退化[J]. 应用生态学报, 1999, 10(6): 743-747.
[5] 刘帅, 高永光. 铜胁迫下玉米叶绿素质量比与光谱反射率关系[J]. 辽宁工程技术大学学报(自然科学版), 2008, 27(1): 125-128.
[6] 白宝璋, 孔祥生, 王玉昆, 等. 植物生理学实验指导[M]. 北京: 中国农业科技出版社, 1996 .
[7] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000: .
[8] 邹琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000: 161-162.
[9] 劉涛, 耿文春, 李丽, 等. 混合盐碱胁迫对两种抗性不同的绣线菊光合特性的影响[J]. 东北农业大学学报, 2009, 40(5): 32-36.
[10] 李永杰, 李吉跃, 方晓娟, 等. 铜胁迫对白蜡幼苗叶绿素含量及光合特性影响[J]. 东北林业大学学报, 2010, 38(6): 35-37.
[11]张自坤, 刘作新, 张颖, 等. 铜胁迫对嫁接和自根黄瓜幼苗光合作用及营养元素吸收的影响[J]. 中国生态农业学报, 2009, 17(1): 135-139.
[12] SHI D C, WANG D L. Effects of various salt-alkaline mixed stresses on Arwurolepidium chinense(Trin.)Kitag[J]. Plant and Soil, 2005, 271: 15-26.
[13] 林义章, 徐磊. 铜污染对高等植物的生理毒害作用研究[J]. 中国生态农业学报, 2007, 15(1): 201-204.
[14] 郭银生. 光环境调控对水稻幼苗和黑豆芽苗菜生长发育的影响[D]. 南京: 南京农业大学, 2011.
[15] 高冠龙, 冯起, 张小由, 等. 植物叶片光合作用的气孔与非气孔限制研究综述[J]. 干旱区研究, 2018, 35(4): 929-937.
[16] 武香. 盐胁迫下植物的渗透调节及其适应性研究[D]. 北京: 中国林业科学研究院, 2012.
[17] 赵佳伟, 李清亚, 路斌, 等. 不同品种北美豆梨对NaCl胁迫的生理响应及耐盐性评价[J]. 植物生理学报, 2019, 55(1): 23-31.
[18] 徐磊. 铜胁迫对小白菜生理生化指标的毒害作用[D]. 福州: 福建农林大学, 2003.
[19] 耿晓东, 周英, 于明华, 等. NaCl胁迫对小黄花菜生长及相关生理指标的影响[J/OL]. 广西植物, (2021-03-12)[2021-04-15]. https://kns.cnki.net/kcms/detail/detail.aspx?FileName=GXZW20210311005&DbName=CAPJ2021.
猜你喜欢
作文周刊·小学二年级版(2022年44期)2022-11-29
作文小学中年级(2021年11期)2021-12-11
中老年保健(2021年6期)2021-08-24
学生天地(2020年13期)2020-08-25
人大建设(2019年7期)2019-11-18
作文小学中年级(2018年5期)2018-06-11
金色少年(奇趣科普)(2017年2期)2017-04-18
饮食保健(2017年5期)2017-03-23
农家科技中旬版(2016年12期)2016-04-16
读写算(中)(2016年9期)2016-02-27
