
梨属植物分类研究进展*
2021-09-12 03:20:40田路明董星光曹玉芬霍宏亮徐家玉
中国果树 2021年9期
田路明,董星光,曹玉芬,张 莹,霍宏亮,齐 丹,徐家玉,刘 超
(中国农业科学院果树研究所,农业农村部园艺作物种质资源利用重点实验室,辽宁兴城 125100)
一般认为梨(Pyrus)起源于第三纪的中国西部或西南部山区,向东向西演化发展,主要分布于欧洲、北非、小亚西亚、伊朗、中亚、中国、朝鲜和日本;从地理分界线来讲,天山和兴都库什山以东分布的梨属植物为东方梨,以西的为西方梨[1]。Pyrus是林奈(Linnaeus)于1753年命名的,而在当时包含梨、苹果和榅桲,后因梨和苹果不能相互嫁接繁殖,Miller 于1768年把苹果从Pyrus中分出,命名Malus(苹果属)。梨属分类的第一项综合研究结果由Decaisne发表,包含23个种,分布于6个地理种群。此后Koehne 根据果实萼片的有无把梨属分为2个种群大区,即PashiaKoehne和AchrasKoehne。Fedorov接受了Koehne的2个大区分类,并增加了2个新的大区即XeropyreniaFed.和ArgyromalonFed.,这2个新的种群大区主要包括几个来自高加索的梨种和杂种。Tuz[2]和Browicz等[3]把土耳其的梨种也归纳入高加索种群下面。Challice等[4]结合生化和形态学特征提出了22 个梨基本种概念。近些年来,仍有不少关于新的梨野生种质命名的报道,如在意大利的西西里发现梨的新种P.sicanorum[5]和P.ciancioi[6],伊朗的梨新种P.ghahremanii、P.giffanica、P.zangezura、P.×takhtadzhianii、P.×medvedevii、P.cordifolia[7-9],土耳其的高加索梨新种P.demetriiKuth.[10]等。目前,关于梨属植物包含多少种还没有确切的数字,植物名录网站(http://www.theplantlist.org/)显示有747个梨种命名,其中69个是被认可的梨种,58个同物异名种,其他命名仍未得到解决;该网站显示公认的13 个中国原产梨种是被认可的,而Pyrus trilocularisD.K.Zang &P.C.Huang(崂山梨)也是一个被认可的梨种,国内该梨分类的相关研究报道并不多。另一个国际植物名录网址(https://www.ipni.org/)也有上千个梨属植物命名。然而,也许受新版命名法规“墨尔本法规”的影响,Christenhusz等[11]最近提出了一项新的对梨属植物重新定义的规则,他提议在梨属下合并一大批以前被广泛承认的种。本文围绕梨属植物分类研究的分歧、植物形态分类研究、结合生化指标的分类研究和结合分子技术的分类研究进行相关综述分析,并结合“墨尔本法规”对梨属植物分类研究提出展望,梨属植物的分类对梨资源保护、评价和利用具有重要意义。
1 梨属植物分类的分歧
自林奈创立植物分类法以来,植物分类工作面临一些困难,如单系类群、并系类群和复系类群的问题,因此相继出现三大分类学派,即表征学派、分支学派和进化学派,三大分类学派均对分类研究作出了贡献,“系统发育命名国际学会”甚至提议以“生物学命名谱系法规”替代用了250多年的林奈分类法[12]。自梨属命名以来,不论从宏观层面,还是微观层面,关于梨属植物分类的分歧一直延续至今。梨属植物曾先被归类于梨亚科的梨亚族,又被归类于桃亚科下苹果族的苹果亚族[1,13],再又被归于苹果亚科的苹果亚族[14]。梨属植物具有配子体自交不亲和系统,依赖于其他亲和的授粉者来维持自然繁衍[15]。梨属植物的形态具有高度可塑性和连续性,由于渐进的形态特征表现和频繁的杂交,给梨属植物的正确分类带来了巨大的挑战。由表1可以看出,1890—1985年Koehne、Maleev、Fedorov、Tuz和Terpó等先后把梨属植物分为不同的大区和亚区[16],在不同大区下区分为不同的梨种[9,17](表2)。
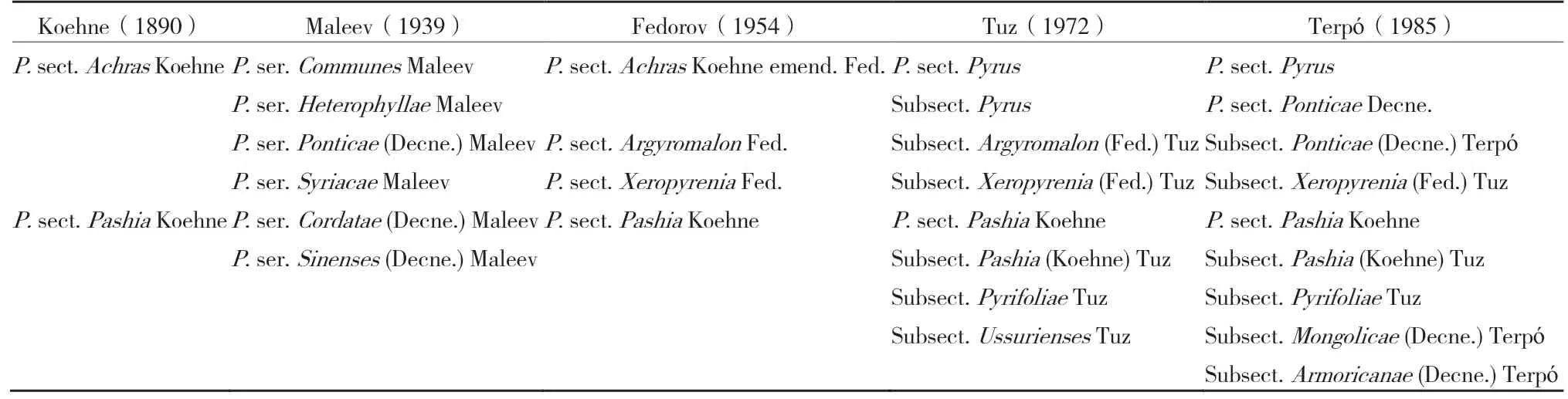
表1 不同年代梨属植物大区分类的比较[16]

表2 伊朗和阿塞拜疆大区分类下梨种情况[9,17]
Rehder 提出了梨属15个基本种,涉及了6个变种和5个近缘种,Challice等[4]对比Rehder 的研究分类,并根据地理分布重新考虑梨属植物的物种命名,描述了22 个基本种。Aldasoro 等[18]研究欧洲和北非的梨种,经与其他专家研究结果进行比较分析,认同了6个种,即P.bourgaeana、P.communis、P.pyraster、P.cordata、P.spinosa和P.nivali。Challice等[4]列出的Kansu pear(甘肃梨)很有可能是新疆梨。据不同报道,梨属的物种数量相差很大,从20种到80 种不等,高加索地区的亚美尼亚约有一半梨属植物类型[16]。我国是东方梨种的主要起源地,我国分类学家相信有13个梨种起源于中国,国外专家Rehder 命名的8个中国梨种P.bretschneideri(白梨)、P.serotina(砂梨)、P.serrulata(麻梨)、P.phaeocarpa(褐梨)、P.betulaefolia(杜梨)、P.calleryana(豆梨)、P.pashia(川梨)和P.koehnei得到中国专家的证实,而其他如P.ovoidea、P.lindleyi和P.kolupana未被发现对应实物存在[19]。中国专家俞德浚命名5个中国梨种P.pseudopashiaYu(滇梨)、P.sikiangensisYu(新疆梨)、P.hopeiensisYu(河北梨)、P.xerophilaY(木梨)、P.armeniacaefoliaYu(杏叶梨)。
2 植物形态分类研究
植物的表型会随着地理环境、光、温、水、气的影响,以及时间的推移而变化,并且个体之间也会有所不同。在梨属分类学研究中,主要依据叶片、果实、枝条和花朵的形态特征,以及枝上有无刺和茸毛的情况,对梨属植物进行分类[20]。然而,刺的存在不能被解释为野性的明确标志,因为刺也存在于已归化的可食用地方梨品种中[21]。P.magyaricaTerpó被描述为匈牙利喀尔巴阡盆地的特有和残留野生物种,Barina 等[22]没有发现与原分类描述相对应植物形态的凭证或活标本,认为P.magyarica应该是P.pyraster,建议停止使用P.magyarica这个名称。
在野生梨分类研究中,一些专家认为叶片特征是重要的分类依据;Terpó[23]认为叶形是梨系统中的主要和限制性特征,由于果实变异性较高,其形态学特征是次要性状,而野生梨有无茸毛不是一个精确的分类依据。格鲁吉亚的野生种P.georgica与土耳其的野生种P.elaeagrifolia形态相似,仅叶尖和叶缘是区分二者的关键特征[24]。以果实为重要分类特征的专家认为,果实萼片的有无、心室数量、果实皮孔和果梗等是重要的分类依据。根据成熟果实萼片的有无,将梨属植物分为2 个大群,其中西方梨大都是萼片宿存,东方梨既有宿萼的,也有脱萼的,然而欧洲野生种P.boisseriana和P.cordata也表现出脱萼现象,其在高加索和中亚也有分布[16]。日本园艺学家菊池秋雄根据果实心室数量将梨属植物分为3个大种群区,即真梨区、豆梨区和杂种区[25]。
Voltas等[21]对野生种P.pyraster和栽培种P.communis的分类研究认为,凭叶片性状不能提供充分的分类依据。在欧洲的3个野生梨种P.pyraster、P.amphigenea和P.nivalis中均发现有3 种类型的果实形状,其中72%为苹果形,楔形占26%,很少为梨形和樱桃形(约1%)[26]。Paganová[20]对斯洛伐克64 个地点的507 份P.pyraster进行评价,根据其叶片和果实特征,将其归为2个亚种P.pyrastersubsp.pyraster和P.pyrastersubsp.achras。
Zywiec等[27]研究了西班牙西部几个不同生态区的野生种Pyrus bourgaeana花性状,发现大部分花朵性状在地理位置上没有差异,然而在不同年份花的性状存在显著差异。Aldasoro 等[18]研究认为,P.bourgaeana和P.communis的花瓣长度分别为5.4~12.0 mm(平均8.7 mm)和12.0~15.0 mm(平均13.2 mm),而Zywiec等[27]调查发现其中约50%的P.bourgaeana花瓣长度大于12 mm。Aydin 等[24]对土耳其的P.pseudosyriaca和P.syriaca的比较分析,前者仅是花梗长度短于后者,认为前者只是P.syriaca的变种P.syriacavar.pseudosyriaca。
3 结合生化指标的分类研究
由于梨属植物形态有相当大的变异,有时只使用植物形态学特征的确很难分离不同种类。Challice等[4]使用29 个化学物质含量如黄酮类和酚类物质,以及22 个叶、果和枝形态指标,编制计算机程序,分析不同梨种之间的分类关系,提出了不同区域的22 个梨基本种。其中P.koehneiSchneider 被认为是中国西南山区和台湾的梨种,但并没有得到中国分类专家的认同,而被认为是豆梨的变种之一[28]。Chouliaras 等[29]利用同工酶LAP 和GOT,对28 个不同梨种质进行聚类分析,第1个组群主要是P.gharbiana、P.balansae、P.lyndleyi及西洋梨品种和砂梨品种,第2个组群主要是P.betulifolia、P.faureii和P.nivalis,第3个组群仅是P.caleryana。林伯年等[30]对梨属的秋子梨、白梨、砂梨、西洋梨、新疆梨、杜梨、木梨、麻梨和褐梨等9 个种的某些品种的过氧化物酶同工酶的测定表明,不同种间谱带差异较大,种内品种间谱带差异较小,西洋梨谱带类型与东方梨有很大差异,白梨和砂梨品种间谱带类型交错,认为白梨和砂梨相似可列为一个种。曹丽等[31]对POD同工酶的研究表明,白梨与砂梨系统品种聚类在一起,将苹果梨归为白梨系统更为合适。马兵钢等[32]利用POD 和EST同工酶的研究表明,苹果梨与秋子梨系统的亲缘关系更近,‘库尔勒香梨’与白梨系统的亲缘关系更近。利用化学成分和同工酶进行分类学研究已经很少了,其对形态分类起到一定的补充验证作用,但仍然存在很多局限性,并不能解释形态差异的根本原因。
4 结合分子技术的分类研究
伴随分子研究的快速发展和分子标记技术的应用,各种分子标记技术被应用于梨属植物的亲缘关系和多样性分析研究。应用于核基因组的分子标记AFLP[33]、SSR[34]等研究,应用于叶绿体基因组的分子标记RFLP[35]、SSR[36]等研究,再到梨基因组重测序的比较研究[37]。梨属植物分子水平的系统树聚类分析研究,对传统的形态学分类结果提供一定的支撑,但是并不能完美解释和解决所有分类难题。
从宏观水平的大量分子研究结果来看,系统树聚类图中东方梨和西方梨都是分开的,栽培品种和野生种质都是分开的。分子技术研究揭示,野生种P.caucasica和P.pyrasterL.Burgsd是欧洲梨栽培品种(P.communisL.)的主要祖先[38],进一步支持了Challice和Westwood 的观点。Bennici等[39]利用SSR分子标记对基因组DNA和叶绿体DNA进行比较分析,表明野生种P.amygdaliformisg更加保守,意大利西西里岛当地梨品种与其他欧洲栽培梨品种相比,与野生种P.pyraster表现出较高的遗传相似性。Zheng 等[40]的研究结果无法最终澄清东亚梨种群是属于单系类群,还是属于欧亚西部种群的并系类群,而其他几种欧亚种(P.elaeagrifoliaPall.、P.nivalisJacq.、P.salicifoliaPall.和P.spinosaForssk.)并非是单系类群。Korotkova 等[16]系统发育重建表明,所采集的高加索梨种群也不是一个单系类群。Wu等[37]对来自世界各地的113份梨种质的基因组重测序分析,包括63份亚洲梨(31 份栽培种和32 份野生种)和50 份欧洲梨(25 份栽培种和25 份野生种),认为梨起源于我国西南地区,经过亚欧大陆进行传播,东方梨与西洋梨分别经历了独立驯化的过程。欧洲唯一栽培梨种(P.communis)来自野生种P.pyraster,亚洲栽培种P.pyrifolia和P.bretschneideri来自共同的祖先野生种P.pyrifolia[36-37],P.sinkiangensis拥有亚洲梨和欧洲梨的遗传背景,而P.regelii、P.armeniacaefolia、P.xerophila、P.hopeiensis和P.fauriei也有混合遗传背景[37]。
从微观水平上的大量分子研究结果来看,分子标记还不能区分芽变品种和部分个体的分类问题。我国的‘库尔勒香梨’和‘苹果梨’的分类也存在争议,形态特征和分子标记各有依据。俞德浚将新疆梨从别的梨种中分离出来,并确立了新疆梨的独立地位,但根据植物形态却将‘库尔勒香梨’归属在白梨系统[41]。分子标记聚类分析表明,既有研究结果支持将‘库尔勒香梨’归属为新疆梨[42],也有研究结果支持将其归属于白梨系统[43]。俞德浚[41]认为从植物形态上‘苹果梨’应归为砂梨系统,而分子标记结果更多支持‘苹果梨’归为白梨系统[34]。Sharifani 等[44]对6 个种34 份梨种质分别进行SSR 分子标记和形态标记聚类分析,结果2个系统树图差异较大,种间交错分布明显,分子标记的遗传距离并不能充分反映形态数据的遗传距离。目前,分子技术还没能结合具体植物形态的局部差异进行分类学鉴定,比如叶尖、叶缘、宿萼、脱萼、有刺和茸毛等。另外根据相关研究,笔者认为如SSR 分子标记引物选择和数量对基因遗传距离的分析也有较大影响。而实际上人工杂交培育的部分新品种或优系种质更是面临分类困难的问题,笔者认为既有物种本身的问题,也有所用分类方法的问题。
5 梨属植物分类的展望
由于单系类群、并系类群和复系类群的存在,不仅梨属植物的分类存在争议难题,其他一些植物等也存在分类的难题,三大分类学派的学术分歧使得林奈分类法的应用面临挑战。历史上由于时间和地理跨度的原因,加之梨属植物种间很容易发生自然杂交,逐渐产生了大量的中间类型,各国专家不能全面掌握相关详细信息,缺乏全面系统的比较分析,给梨属植物的正确分类带来了困难,甚至产生许多同物异名种现象(表3)。另外,生化标记和分子标记并不能完美对应解释植物形态的差异,目前还不足以解决以植物形态差异进行分类所面临的诸多问题。

表3 梨属分类中的部分同物异名种及变种
近些年来,由于计算机技术的飞速发展,人工智能已经应用于诸多领域,图像识别技术将有可能辅助解决植物分类的部分难题。《自然》杂志报道,研究人员利用扫描植物标本卡获得的2.6 万余幅图像对类似算法进行了训练,该计算机程序最终能以近80%的准确度鉴别出植物物种,这可能比分类学家的表现好很多[45]。翟果等[46]利用数字图像处理技术识别观赏菊品种的方法,结果表明20 个品种的平均正确识别率可达到92.17%,验证了利用图像处理技术识别观赏菊品种的可行性。张雪芹等[47]利用基于深度学习的图像识别算法,训练所得的植物识别模型可识别206种植物,准确率达到86.7%,并以此模型为基础开发了包含多个应用前端的智能植物图像识别系统供测试和使用。
2011年在澳大利亚墨尔本召开的第十八届国际植物学大会表决通过了“墨尔本法规”,其中几个重要的变化将影响新名称的发表,提供了有关电子出版物的命名新规则的草案文本,并概述了相应的最佳做法;今后新名称的作者、编辑和出版商都将确保含新名称的出版物符合“墨尔本法规”,以便其中的名称是有效发表[48]。结合新的相关命名法规“墨尔本法规”,应加强国际间的合作研究,通过组织召开关于梨属植物分类学国际大会,统一规范对梨属植物的分类标准,建立更加丰富的梨属植物分类信息互联网数据库,借助人工智能图像识别技术和分子技术,也许能让计算机替代人工有效解决梨属植物的分类问题。
猜你喜欢
作物学报(2022年2期)2022-11-06 12:11:02
中国烟草学报(2021年3期)2021-08-04 03:50:42
农村青少年科学探究(2020年5期)2020-08-18 02:20:52
东坡赤壁诗词(2019年3期)2019-07-05 06:55:54
新民周刊(2018年8期)2018-03-02 15:45:54
江苏农业科学(2017年15期)2018-02-06 07:17:42
饮食科学(2017年12期)2018-01-02 09:23:20
上海农业学报(2017年3期)2017-04-10 12:39:10
爱你(2016年18期)2016-11-25 19:31:30
课堂内外·创新作文高中版(2016年10期)2016-11-08 06:06:30
