
不同冬季作物对稻田蚯蚓及蚯蚓粪产量的影响*
2021-09-10 08:18:38郭立君程凯凯唐海明汤文光肖小平
中国生态农业学报(中英文) 2021年9期
李 超, 赵 杨, 郭立君, 程凯凯, 唐海明, 黄 敏,汤文光, 文 丽, 汪 柯, 崔 婷, 肖小平**
(1.湖南省土壤肥料研究所 长沙 410125; 2.湖南农业大学作物环境研究中心 长沙 410128;3.湖南省水稻研究所 长沙 410125; 4.南县农业局 南县 413200)
蚯蚓营养丰富, 蛋白质含量占干重的56%~65%,蚯蚓自溶液的游离氨基酸含量高达50~70 g∙L−2, 含有10种人体必需的氨基酸, 不饱和脂肪酸含量高,饱和脂肪酸含量低, 微量元素含量丰富, 并因其特殊的生活环境促使体内含有多种活性成分[1]。蚯蚓除可作为传统的中药材原料外, 还可用于提取“蚓激酶”“氨基酸”作为轻工业的重要原料, 以及为现代畜牧业、渔业提供优质的饲料及饵料[2]。因此, 目前市场上的蚯蚓缺口较大, 蚯蚓养殖将是大势所趋, 但蚯蚓人工养殖存在劳动力投入大、养殖成本高及及大规模养殖难度大等难题。中国南方稻区大约有9.3×106hm2冬闲田, 其中72%较容易开发利用[3]。冬季的低温高湿气候及冬季作物的旱作环境为蚯蚓提供了良好的生存条件; 同时, 冬季作物的落叶及前茬的稻草秸秆可为蚯蚓提供充足的食物及适宜的生长繁殖条件。因此, 通过在稻田种植冬季作物, 实行水旱轮作, 充分利用冬季的温光水热资源, 挖掘稻田蚯蚓产量潜力, 有利于缓解蚯蚓市场缺口大这一难题。
目前, 在稻田水旱轮作系统仅在小麦(Triticum aestivum)-水稻(Oryza sativa)、油菜(Brassica napus)-水稻系统有过少量关于蚯蚓的报道[4-7], 主要集中在蚯蚓提高水稻产量及稻米品质[4-5,8], 改善土壤结构[9],活化土壤养分[10-13], 优化土壤微生物群落结构[14]等方面, 而有关冬季作物对稻田蚯蚓及蚯蚓粪产量的影响研究尚少见相关报道。落叶是蚯蚓的重要食物来源。通常来说, 作物落叶生物量伴随作物生物量的增加而增加[15-16], 且不同作物落叶的C/N比不同, 蚯蚓更青睐于取食低C/N比的有机物料[17-19]。同一作物在栽培措施一致的条件下, 高地力水平下良好的土壤环境有利于作物高产[20], 其较高的落叶生物量可保障蚯蚓食物的充足供应; 同时, 高地力水平下的土壤有机质含量较高, 其有利于蚯蚓种群的增长[21], 而蚯蚓产生的蚯蚓粪可显著增加后茬作物的地上部生物量及产量[22], 有利于促进稻田水旱轮作系统的可持续性发展。因此, 本研究基于大田调查, 以冬闲田为对照, 拟开展冬季种植紫云英(Astragalus sinicus)及油菜不同产量水平下的稻田蚯蚓及蚯蚓粪产量差异研究, 以期为促进稻田蚯蚓增效及缓解蚯蚓市场缺口提供一定理论及技术支撑。
1 研究区域与研究方法
1.1 研究区域与供试材料
研究区域地处湖南省益阳市南县三仙湖镇, 年平均气温16.6 ℃, 降水量1237.7 mm, 雨日136.3 d, 日照时数1775.7 h, 蒸发量1236.2 mm。属亚热带过渡到季风湿润气候类型。土壤类型为紫潮泥黏土。该区域采用水旱轮作模式, 淹水季种植水稻, 旱作季种植冬季作物紫云英及油菜, 品种分别为‘湘紫1号’和‘华湘油16号’, 均采用免耕直播栽培。
蚯蚓鉴定主要参照《中国动物图谱——环节动物》[23]和《中国土壤动物检索图鉴》[24]。经鉴定,该研究区域的优势蚯蚓种群为土壤中自然存在的深栖类表土排粪的威廉环毛蚓(Pheretima guillelmi)[4],其在冬季作物生长季由田埂等周边生境陆续迁至田间进行生长繁殖, 水稻生长季灌水后由田间迁至田埂等周边生境进行避难过渡。因此, 本研究仅研究冬季作物生长期内的蚯蚓(威廉环毛蚓)及蚯蚓粪产量。
1.2 研究设计
同一区域同一土壤类型下, 稻田不同冬季作物不同产量水平下的落叶类型、落叶产量等差异较大,进而可能影响蚯蚓的生长繁殖。因此, 为探明自然生产条件下不同冬季作物不同产量水平间的蚯蚓及蚯蚓粪产量差异, 本研究基于2017−2020年的大田调查, 以冬闲(T1)为对照, 将冬种紫云英(T2)及油菜(T3)的不同田块的产量划分为低、中、高3类,分别为低产紫云英(T2-L)、中产紫云英(T2-M)、高产紫云英(T2-G)、低产油菜(T3-L)、中产油菜(T3-M)、高产油菜(T3-G), 共计7个调查处理。每个调查处理挑选有代表性的田块5块, 每个田块面积2000 m2左右。种植冬季作物的田块在水稻收获后,及时用开沟机开好厢沟、围沟和腰沟, 沟深15~20 cm,沟宽20~30 cm, 做到沟沟相通。紫云英10月中旬播种, 播种量为22.5 kg∙hm−2, 油菜10月中旬播种, 采用免耕直播栽培, 播种、施肥及病虫草害管理参照作者已发表文章[25]。
1.3 测定项目与方法
降雨量及气温: 数据来源于湖南省益阳市南县气象局, 包括2017−2020年冬季作物生长期(10月至翌年4月)的逐日平均气温、逐日降雨量。5 ℃为蚯蚓的生物学起点温度, 低于5 ℃蚯蚓进入休眠状态。故活动积温Y=N×T,N为气温≥5 ℃的天数,T为气温≥5 ℃的平均气温; 有效积温K=N(T−5)。
土壤有机质: 2020年油菜成熟期及紫云英翻压前, 每个调查田块用土钻按棋盘法选取15个点, 取0~20 cm耕层土壤, 室内风干除杂后, 采用高温外加热重铬酸钾氧化-容量法测量土壤有机质含量。
冬季作物测产: 2020年油菜成熟期, 每个田块按5点取样法选取5个取样点, 每个取样点测量4 m2,脱粒风干后称取干重, 然后以9%的籽粒含水量计算油菜产量。紫云英盛花期, 挑晴朗天气在每个田块选取5个取样点, 每个取样点测量1 m2, 称鲜重测产。
蚯蚓及蚯蚓粪测产: 在油菜及紫云英测产后, 对应每个田块按对角线取样法, 每条对角线取样7个点, 共13个点, 每个点测量1 m2, 取样深度20 cm, 将蚯蚓人工挖出并计数, 去除泥土等杂物后称鲜重。同时在每个调查点内选取40 cm×30 cm的代表性样方, 将其土壤表层蚯蚓粪收集带回实验室, 去除秸秆、福寿螺壳等杂物后风干称重。最后按0.9的厢面系数(厢宽∶沟宽=9∶1)折算冬季作物生长期内的蚯蚓及蚯蚓粪产量。此处, 水稻生长季灌溉阶段(5月下旬至9月中旬), 蚯蚓无法生存, 水稻生育后期(9月下旬至10月上旬)断水后, 虽有幼龄蚯蚓逐渐迁入田埂附近区域, 但蚯蚓粪产量极低, 可忽略。因此,本文中的蚯蚓粪产量被视为冬季作物生长期内蚯蚓所产生的粪的产量。

变幅为T2或T3模式下3个冬季作物产量水平(L、M、G)各指标值的最大值减去最小值再除以平均值。
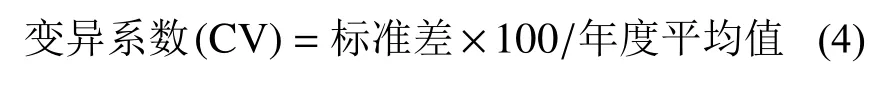
1.4 数据处理
运用SPSS Statistics 21、Excel 2007实用数据分析软件对试验数据进行统计分析和作图。
2 结果与分析
2.1 试验期间降雨量和气温的变化特征
表1表明: 不同年度间冬季作物生长期内的降雨量差异较大, 2018−2019年10−12月的降雨量占总降雨量的43.1%, 而2017−2018年、2019−2020年仅分别为21.7%、21.3%。2018−2019年10月、11月、12月的降雨量分别为2017−2018年的1.9倍、1.9倍和9.0倍, 为2019−2020年的1.9倍、2.3倍和3.1倍。降雨量CV值较大的月份主要集中在10月至翌年2月, 在32.6%~97.5%波动, 表现为12月>11月>10月>1月>2月。结果表明10−12月是造成不同年度间冬季作物生长期内降雨量差异的关键月份。

表1 2017—2020年试验区冬季作物生育期内的降雨量Table 1 Rainfall during the growing season of winter crops in 2017−2020 in the study area
表2表明: 冬季作物生长期(10月至翌年4月)内, 连续3年气温≤5 ℃的月份均出现在12月至翌年1月, 且不同年度间的霜冻(≤0 ℃)天数波动较大。≤5 ℃及霜冻期主要集中在1月。不同年份间冬季作物生长期内的活动积温差异较小, CV值仅为7.3%, 但12月至翌年2月的活动积温CV值达31.0%, 其中2018−2019年的活动积温最低, 仅为353.5 ℃, 较2017−2018年、2019−2020年分别降低39.8%、47.8%。2018−2019年12月、1月、2月的活动积温较2017−2018年分别降低34.5%、25.1%、54.4%, 较2019−2020年分别降低38.5%、30.5%、64.9%。不同年度间及不同月份间的有效积温差异的表现规律同活动积温一致。表明不同年份间冬季作物生长期内的积温差异较大, 主要集中在12月至翌年2月的低温期。

表2 2017—2020年研究区冬季作物生育期内的积温Table 2 Accumulated temperature during the growing season of winter crops in 2017−2020 in the study area
2.2 冬季作物产量
2019年紫云英(T2)平均产量较2018年和2020年分别减少33.5%和37.3%, 2019年油菜(T3)平均产量较2018年和2020年分别减少20.3%和36.5%。2019年的紫云英产量变幅较2018年、2020年分别增加15.0个百分点、23.4个百分点, 油菜分别增加5.8个百分点、30.5个百分点, T2的平均变幅较T3增加63.1个百分点。紫云英的产量CV值变化范围为19.0%~30.2%, 表现为T2-M>T2-L>T2-G。油菜产量CV值变化范围为17.9%~37.9%, 表现为T3-L>T3-M>T3-G。油菜产量CV值变化范围大于T2, 但T3的平均CV值较T2仅增加2.4%, 未达显著差异(表3)。结果表明同一冬季作物不同年份间的产量变幅差异较大, 高产田块的冬季作物产量变异小, 稳产性更好, 以冬种油菜效果最佳。
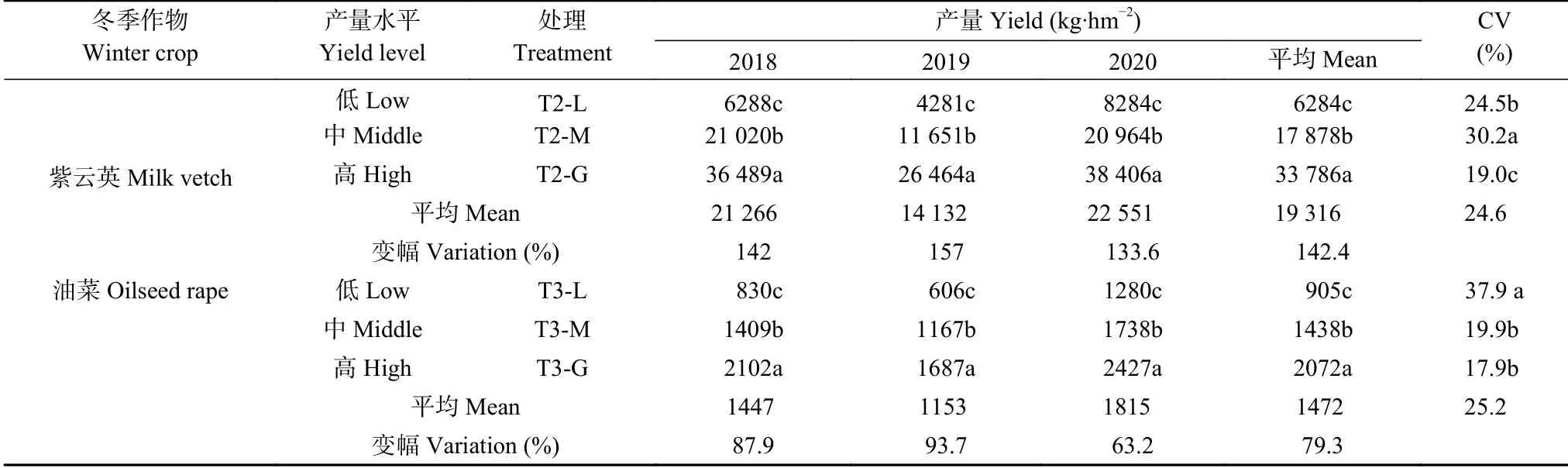
表3 2018—2020年冬季作物产量Table 3 Yields of winter crops in 2018−2020
2.3 土壤有机质
土壤有机质含量(3种产量水平的均值)表现为T3>T2>T1, 冬种紫云英及油菜的土壤有机质含量较冬闲分别增加10.1%、16.0%(图1)。冬种紫云英及油菜条件下, 土壤有机质含量均伴随冬季作物产量的增加而显著递增(P<0.05)。表明种植冬季作物有利于提高土壤的有机质含量, 其中冬种油菜效果更好。
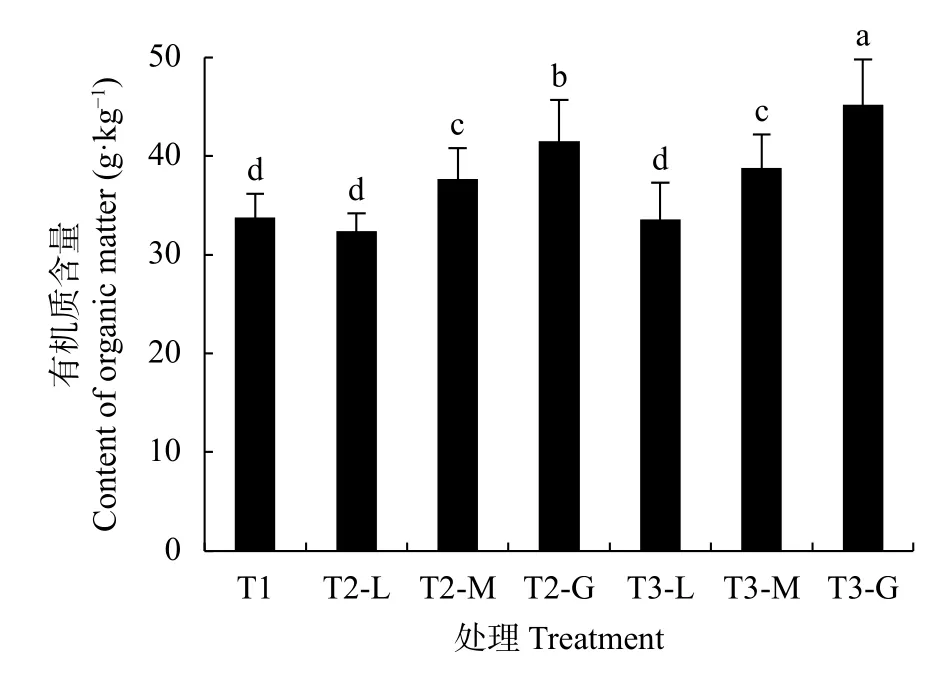
图1 不同冬季作物条件下的土壤有机质含量Fig.1 Effects of different winter crops and their yield levels on the content of soil organic matter
2.4 蚯蚓产量
从表4可知, T1的蚯蚓密度及蚯蚓产量范围分别为8.4~12.8条∙m−2和290~428 kg∙hm−2, T2分别为11.2~30.8条∙m−2和368~1054 kg∙hm−2, T3分 别 为20.4~49.1条∙m−2和763~1845 kg∙hm−2, 平均蚯蚓密度及蚯蚓产量均表现出T3>T2>T1, T2和T3的平均蚯蚓密度较T1分别显著增加91.7%和236.1% (P<0.05),T2和T3的平均蚯蚓产量较T1分别显著增加86.2%和265.2%, T3的平均蚯蚓密度及蚯蚓产量较T2分别显著增加75.4%和96.1%。同时, 蚯蚓密度及产量在不同年度间均表现出较大的波动。冬种紫云(T2)条件下, 密度CV值伴随紫云英产量的增加呈现先增后降的趋势, 而冬种油菜(T3)条件下, 则伴随油菜产量的增加而降低, 但均以T2-G和T3-G的蚯蚓密度CV值最低。蚯蚓个体生物量表现为T3>T1>T2, T3的个体生物量较T2、T1分别增加11.9%和8.7%。T2、T3不同年度间的平均蚯蚓密度及产量均伴随冬季作物产量的增加而增加, 且T2不同年度间的平均蚯蚓密度及产量变幅大于T3, 较T3分别增加17.1个百分点及17.0个百分点。表明冬季作物高产的田块, 其蚯蚓产量与产量稳产性也较高, 以冬种油菜效果更好。

表4 不同冬季作物条件对蚯蚓密度、产量及个体生物量的影响Table 4 Effects of different winter crops and their yield levels on earthworm density, yield, and individual biomass
2.5 蚯蚓粪产量
总体上看, 蚯蚓粪产量表现出同蚯蚓产量相似的趋势(表5)。T1、T2、T3的蚯蚓粪产量范围分别为1921~3126 kg∙hm−2、3023~10 116 kg∙hm−2和6034~18 958 kg∙hm−2, 个体蚯蚓粪产量范围分别为22.9~25.8 g∙条−1、27.0~33.8 g∙条−1和29.6~39.2 g∙条−1, 均表现出T3>T2>T1。T2、T3的平均蚯蚓粪产量较T1分别显著增加142.7%和373.0%, 平均蚯蚓粪个体产量分别显著增加25.2%和38.9%。T2、T3不同产量水平间的蚯蚓粪产量在不同年度间的波动规律同前文的蚯蚓产量。T3的平均蚯蚓粪产量及个体产量较T2分别显著增加94.9%和11.0%。T2、T3不同年度间的平均蚯蚓粪产量及个体产量均伴随冬季作物产量的增加而增加, 且T2不同年度间的平均蚯蚓粪产量变幅较T3增加14.6个百分点, 而蚯蚓粪个体产量变幅较T3仅降低1.6个百分点。表明冬季作物高产的田块, 其蚯蚓粪产量与产量稳产性也较高, 冬种油菜效果更好。

表5 不同冬季作物对蚯蚓粪产量及个体产量的影响Table 5 Effects of different winter crops and their yield levels on yield of earthworm cast
2.6 粪蚓比
T1、T2和T3不同产量水平下的平均粪蚓比分别为7.1、9.1和9.0, 平均为8.8, 说明稻田每1 kg威廉环毛蚯蚓可产生8.8 kg蚯蚓粪。T1的粪蚓比显著低于T2、T3, T2、T3不同产量水平下的平均粪蚓比相对T1分别增加28.2%和26.8%, T2相对T3仅增加1.1%, 未达显著差异。同一模式下, 粪蚓比伴随冬季作物产量的增加而增加, T2、T3不同产量水平下的粪蚓比变幅表现为T3>T2, 分别为11.6%和14.3%。表明种植冬季作物可显著提高粪蚓比, 且冬季作物高产田块的粪蚓比更高(图2)。
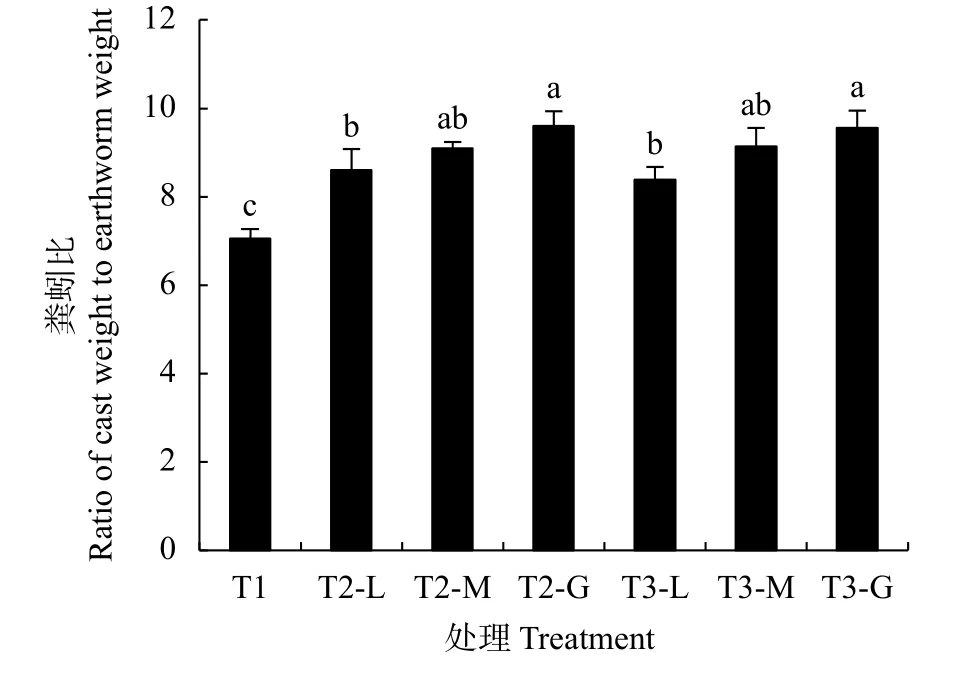
图2 不同冬季作物对粪蚓比的影响Fig.2 Effects of different winter crops and their yield levels on the ratio of cast weight to earthworm weight
2.7 蚯蚓产量、蚯蚓粪产量与作物产量、土壤有机质的相关分析
蚯蚓产量、蚯蚓粪产量与土壤有机质含量均显著(P<0.05)正相关(图3a, d), 与紫云英鲜草产量及油菜产量均呈极显著(P<0.01)正相关(图3b, c, e, f)。同时, 前文已证实不同年度间冬季作物生长期内的降雨及积温差异较大, 导致不同年度间的蚯蚓产量、蚯蚓粪产量及冬季作物产量波动较大, 且蚯蚓及蚯蚓粪不同年度间的产量CV值与冬季作物产量CV值极显著正相关(图4)。因此, 冬季作物高产田块,其土壤有机质含量较高, 有利于提高蚯蚓产量、蚯蚓粪产量及其产量稳定性。
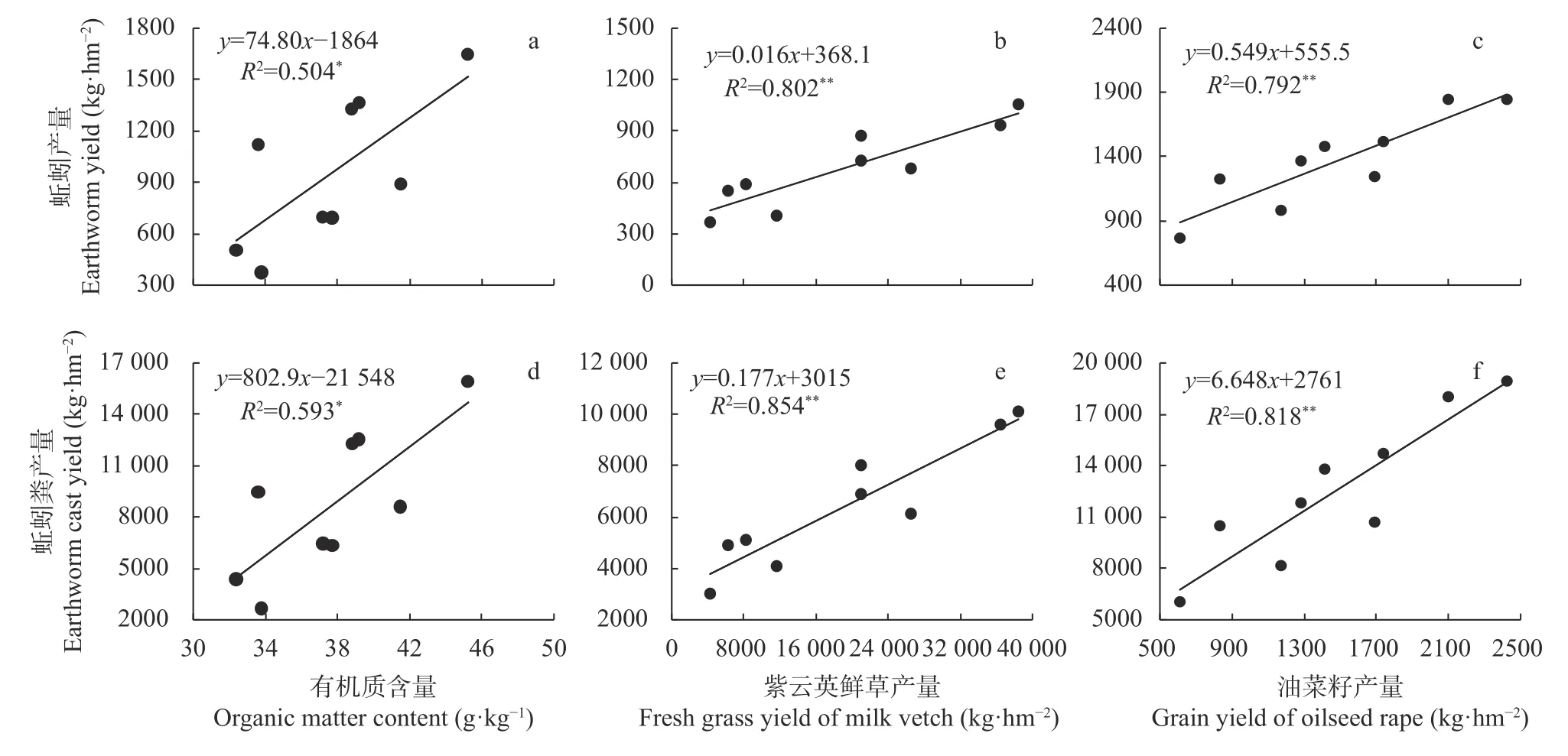
图3 蚯蚓产量(a, b, c)及蚯蚓粪产量(d, e, f)与土壤有机质和冬季作物紫云英、油菜产量的相关系Fig.3 Correlations of earthworm yield (a, b, c) and earthworm cast yield (d, e, f) with soil organic matter content and yields of winter crop of milk vetch and oilseed rape

图4 冬季作物产量变异系数(CV)与蚯蚓产量及蚯蚓粪产量变异系数(CV)间的相关性Fig.4 Correlations of coefficients of variation (CV) of earthworm yield, earthworm cast yield with winter crop yield
3 讨论
冬季作物易受持续阴雨及低温冻害等不利气候的影响[26]。如油菜产量与苗期日平均气温呈显著正相关, 与全生育期降雨量呈负相关, 其中冬前苗期持续降雨导致的减产风险最大[26-27]。蚯蚓属中温型湿生动物, 对土壤温度和湿度变化较为敏感, 蚯蚓生长繁殖的最适温度为20~25 ℃, 最适湿度为35%~60%,且土壤湿度对幼蚓孵化的影响明显强于土壤温度[28-29]。本研究表明, 不同月份在2017−2020年的降雨量及积温CV值的波动幅度均较大, 表现为降雨量大于积温, 且降雨量及积温差异最大的月份分别集中在10−12月、12−2月, 2018−2019年10−12月的持续降雨及12−2月的持续低温导致了蚯蚓的大幅减产, 这与刘满强等[30]研究证实的蚯蚓种群的季节波动性强的研究结果一致。这主要由于10−12月的强降雨易导致免耕稻田渍水, 抑制了蚯蚓种群的增长, 降低了稻田蚯蚓前期繁殖的基数。同时渍水易导致油菜等冬季作物烂根、叶片功能锐减,植株水分亏缺严重, 渍害消除后恢复生长慢, 生物量大幅下降[31-32], 导致可供蚯蚓取食的落叶数量大幅减少, 从而进一步抑制了蚯蚓种群的增长, 这在本研究中冬季作物的产量CV值与蚯蚓产量CV值呈极显著正相关的结果中进一步得到验证。
与冬闲相比, 种植冬季作物可显著增加蚯蚓的密度及产量[4,33]。Huang等[4]研究发现, 冬种油菜较冬闲可显著增加蚯蚓(Pheretima guillelmi)密度68%~154%。本研究表明: 冬种紫云英、油菜的蚯蚓密度较冬闲分别显著增加91.7%和236.1%, 蚯蚓产量较冬闲分别显著增加86.2%和265.2%, 这可能主要由于冬种油菜或紫云英条件下的落叶为蚯蚓提供了充足的食物, 从而促进种群的增长。同时, 覆盖植被类型决定了归还土壤的有机物数量和质量, 也是影响蚯蚓种群分异的主要驱动因子[30,34]。蚯蚓更青睐于取食低C/N比的有机物料, 如种植豆科作物更有利于促进蚯蚓种群密度及产量的增加[33,35]。Hubbard等[19]证实, 与小麦-玉米(Zea mays)轮作相比, 大豆(Glycine max)-玉米轮作可显著增加蚯蚓(Aporrectodea trapezoides)密度96.8%, 产量增加31.5%。Bilalis等[36]证实, 相比棉花(Gossypiumspp.), 豆科作物更有利于蚯蚓种群的增长。但本研究表明: 冬种油菜条件下的蚯蚓密度、蚯蚓产量及蚯蚓粪产量均显著高于豆科植物紫云英, 可能是因为冬种油菜条件下的土壤有机质含量高于紫云英, 而高有机质含量的土壤更有利于蚯蚓种群的生长繁殖。同时, 也可能与蚯蚓更喜欢取食油菜落叶有关。
通常来说, 在相同的外界环境条件下, 作物生物量与作物产量呈正相关关系[37]。因此, 高产田块的作物秸秆或落叶的数量高于中、低产田块, 从而增加了可供蚯蚓取食的食物来源, 有利于蚯蚓种群的生长繁殖。本研究表明: 免耕条件下蚯蚓密度、蚯蚓产量、蚯蚓粪产量及粪蚓比均伴随紫云英及油菜产量的增加而显著增加, 这表明蚯蚓产量与冬季作物产量是协同互促的关系, 冬季作物高产田块的土壤环境有利于增加蚯蚓种群, 促进免耕条件下土壤表层蚯蚓粪的积累。本研究中蚯蚓每年可产蚯蚓粪1.9~11.9 t∙hm−2, 这低于竹林的42.6 t∙hm−2[38], 玉米及木薯(Manihot esculenta)系统中的15.2~432.0 t∙hm−2[39],这可能主要是因为气候及作物系统的差异所致。土壤表层的蚯蚓粪主要功能有: ①减少土壤蒸发, 蒸发量伴随蚯蚓粪覆盖度和厚度的增加而减小[40]。②增加土壤有机质含量, 新鲜蚯蚓粪的有机质含量可达土壤有机质含量的3.5倍, 有机质能够促进团聚体结构的形成, 增加土壤孔隙, 增强土壤保水性能[40-42]。③加速土壤养分矿化及循环[43]。④增加有益微生物种群, 优化微生物群落结构[14]。蚯蚓粪通过以上功能直接或间接地促进作物根系生长及地上部生物量的积累, 提高作物产量[22,44], 最终实现作物产量与蚯蚓产量的协同提高。
本研究只基于大田调查开展了不同冬季作物对蚯蚓及蚯蚓粪产量的初步探索, 年限较短, 应开展长期跟踪研究, 进一步明确气候、冬季作物产量及蚯蚓产量三者之间的关系。同时, 下一步应开展室内盆栽实验及大田小区试验, 就冬季作物落叶数量及质量对蚯蚓生长繁殖的影响进一步开展相关研究。
4 结论
冬季作物、蚯蚓产量及蚯蚓粪产量在不同年度间的波动幅度均较大, 10−12月的持续降雨及12月至翌年2月的低温是导致2018−2019年冬季作物及蚯蚓产量大幅下降的关键因素。蚯蚓产量、蚯蚓粪产量与土壤有机质含量显著正相关, 与冬季作物产量均呈极显著正相关。蚯蚓密度、蚯蚓产量及蚯蚓粪产量均表现为冬种油菜>冬种紫云英>冬闲, 且油菜不同产量水平间的蚯蚓产量及蚯蚓粪产量的变幅均小于紫云英。粪蚓比伴随冬季作物产量的增加而增加, 平均为8.8。综上, 蚯蚓产量及蚯蚓粪产量受不同年际间的气候干扰较大, 冬季作物高产田块的土壤有机质含量较高, 有利于提高蚯蚓产量、蚯蚓粪产量及其产量稳定性, 且冬种油菜的效果更好。
猜你喜欢
诗歌月刊(2024年1期)2024-02-05 01:36:51
土壤学报(2022年1期)2022-03-08 08:36:10
中学时代(2018年6期)2018-11-20 05:53:20
海峡姐妹(2018年7期)2018-07-27 02:30:16
作文小学中年级(2018年5期)2018-06-11 01:16:12
小学生导刊(2017年19期)2017-07-19 13:41:50
快乐语文(2017年9期)2017-06-15 20:28:36
生物灾害科学(2015年3期)2015-03-11 16:38:42
湖南农业科学(2014年24期)2014-02-27 14:33:39
湖南农业科学(2014年20期)2014-02-27 14:32:56
